

1.Возбудимые ткани – это нервная,мышечная и железистая структуры, которые способны спонтанно или в ответ на действие раздражителя возбуждаться.
*Нервная ткань состоит из нервных клеток – нейронов и клеток нейроглии. Кроме того, она содержит рецепторные клетки.
*Мышечная ткань-способна к сокращению,что обеспечивает двигательную функцию органов и организма.. Во всех клетках мышечных тканей содержатся тончайшие сократительные волоконца — миофибриллы, образованные линейными молекулами белков — актином и миозином. При скольжении их относительно друг друга происходит изменение длины мышечных клеток.
*Поперечнополосатая (скелетная) мышечная ткань построена из множества многоядерных волокноподобных клеток длиной 1—12 см.. Из нее построены все скелетные мышцы, мышцы языка, стенок ротовой полости, глотки, гортани, верхней части пищевода, мимические, диафрагма. Особенности поперечнополосатой мышечной ткани: быстрота и произвольность (т. е. зависимость сокращении от воли, желания человека), потребление большого количества энергии и кислорода, быстрая утомляемость.
*Сердечная ткань состоит из поперечно исчерченных одноядерных мышечных клеток, но обладает иными свойствами. Клетки расположены не параллельным пучком, как скелетные, а ветвятся, образуя единую сеть. Благодаря множеству клеточных контактов поступающий нервный импульс передается от одной клетки к другой, обеспечивая одновременное сокращение, а затем расслабление сердечной мышцы, что позволяет ей выполнять насосную функцию. Клетки гладкой мышечной ткани не имеют поперечной исчерченности, они веретеновидные, одноядерные, их длина около 0,1 мм. Этот вид ткани участвует в образовании стенок трубкообразных внутренних органов и сосудов (пищеварительного тракта, матки, мочевого пузыря, кровеносных и лимфатических сосудов). железистый эпителий — образует большинство желез (щитовидную, потовые, печень и др.), клетки которых вырабатывают тот или иной секрет.
*Эпителиальные ткани имеют следующие особенности: их клетки тесно прилегают друг к другу, образуя пласт, межклеточного вещества очень мало; клетки обладают способностью к восстановлению (регенерации). Эпителиальные клетки по форме могут быть плоскими, цилиндрическими, кубическими. По количеству пластов эпителии бывают однослойные и многослойные.
Раздражимость- способность тканей изменять обмен веществ и отвечать на действие раздражителей.
Возбудимость —способность клеток отвечать на раздражение возбуждением. К возбудимым относят нервные, мышечные и некоторые секреторные клетки.
Раздражение – возникаети под действием разл.факторов,кот. Вызывают ответную реакцию. (Раздражители бывают разл.видов. Вопрос №3.)
Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения).
Проводимость- способность проводить возбуждение,т.е. проводить ПД. (скорость проведения у нерва-до 120м/c) Возбуждение и раздражение отличаются: раздражение-специфическая реакция, возбуждение распространяется по организму и является более кратковременной реакцией.
2.Фосфолипиды образуют двойной прерывистый слой. В этот слой включены белки, полярные группы которых сохраняют контакт с водной фазой. Некоторые белки пронизывают мембрану насквозь, другие погружены в липидный бислой наполовину. Часть белков связана друг с другом; другие в большей или меньшей степени окружены липидами. Одни из них являются ионными каналами, другие содержат боковые цепи гликозаминогликанов. Поверхность мембраны покрыта гликокаликсом — трехмерной сетью нитей гликозаминогликанов, соединенных между собой при помощи кальциевых мостиков. Гликокаликс обеспечивает механическую прочность мембраны, участвует в межклеточных взаимодействиях, рецепции, иммунологическом дифференцировании, разделяет молекулы веществ, контактирующих с клеткой, по величине и заряду. Молекулы липидов, образующих бислой, амфотерны. Своими гидрофильными головками они обращены в сторону водных фаз (межклеточная жидкость и цитоплазма) и формируют внешнюю и внутреннюю поверхности мембраны. Важнейшей особенностью мембранных липидов является способность к перекисному окислению (ПОЛ) с образованием свободных радикалов. Функциональное отличие мембраны одной клетки от мембраны другой определяется наличием в ней специфических мембранных белков.Белки, погруженные в фосфолипидный слой и пронизывающие его насквозь, называются внутренними мембранными белками, или белковыми каналами.Другие белки — периферические — прикреплены к поверхности клетки. С учетом выполняемых функций мембранные белки всех клеток делят на 5 классов: белки-насосы, белки-каналы, белки-рецепторы, ферменты и структурные белки.
Важнейшими функциями клеточных мембран являются барьерная, биотрансформирующая, транспортная, рецепторная, генерация электрических потенциалов и образование межклеточных контактов.
Белки-каналы представляют собой пути избирательного переноса ионов и заряженных молекул. Механизм переноса связан с конформацией белка-канала, в результате которой он открывается или закрывается. Взаимодействие рецептора с соответствующим ему лигандом инициирует закрытие или открытие связанного с рецептором канала.
Ионоселективные каналы делят на химические и электрозависимые. В первом случае раздражителем является вещество (медиатор, гормон, метаболит, лекарственное средство), во втором — возникающее в непосредственной близости от электрозависимого канала возбуждение, т.е. потенциал действия.
Ионоселективные каналы в зависимости от скорости их активации и переноса ионов делят на быстрые (например, натриевые) и медленные (например, калиевые, кальциевые).
Для каждого из переносимых через мембрану вида ионов существуют самостоятельные транспортные системы — ионные каналы (натриевые, калиевые, кальциевые, каналы для хлора), основные свойства и механизмы действия которых сходны. Ионный канал состоит из поры, воротного механизма, сенсора (индикатора), напряжения ионов в самой мембране и селективного фильтра.
3.Раздражителями, вызывающими возбуждение, являются различные внешние (действующие из окружающей среды) и внутренние (возникающие в самом организме) воздействия. Все они различны по виду энергии!
Внешние: 1) Физические- механические (щипок,удар), электрические( самый близкий,т.к. это нервный импульс), термические, звуковые, лучевые (свет, рентгеновские лучи и др.) воздействия. 2) Химические -пищевые, вещества, поступающие в организм, кислоты, щелочи и другие\раздражающие жидкости, яды и лекарственные вещества. 3)Биологическими -вирусы, микроорганизмы, насекомые и другие живые организмы. 4) Социальный раздражитель- ятрогенный врач Внутренние- физико-химические раздражители (изменение осмотического и онкотического давления,pH крови, коллоидного состояния, различные физиологически активные вещества, образующиеся в организме и изменяющие деятельность его органов (гормоны, продукты обмена веществ, например углекислота и др.).
По физиологическому раздражению адекватные (специфические) и неадекватные (неспецифические). К адекватным -раздражители, которые влияют в естественных условиях на возбудимые системы организма, приспособленные в процессе эволюции к восприятию этих воздействий. Реакцию вызывают при очень высокой силе.(для органа зрения адекватным раздражителем является свет, для органа слуха — звуковые волны, для температурных рецепторов кожи— тепло и холод )Неадекватными являются такие раздражители, к восприятию которых возбудимые системы биологически не приспособлены. При значительной силе подобных раздражителей и они могут вызвать возбуждение в данных системах. Так, мышца может сокращаться под влиянием механического, теплового и других воздействий, хотя естественными раздражителями для нее являются нервный импульс и некоторые физиологически активные вещества. Чтобы вызвать возбуждение, неадекватный раздражитель должен 'быть во много раз сильнее, чем адекватный для данного воспринимающего аппарата.
4. Реакция клеток, тканей на раздражитель определяется законами раздражения:
1.Закон "все или ничего"-это зависимость амплитуды ответа возбудимой ткани от силы раздражителя .При подпороговых раздражениях клетки, ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца.
2.Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость.
3.Закон силы-длительности.(времени) Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы-длительности. По этой кривой можно определить ряд параметров возбудимости.
а) Порог раздражения – это минимальная сила раздражителя, при которой возникает возбуждение.
б) Реобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани.
в) Полезное время – это минимальное время действия раздражителя силой в одну реобазу, за которое возникает возбуждение.
г) Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возникновения возбуждения. Этот параметр предложил рассчитывать Л. Лапик, для более точного определения показателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбудимость и наоборот.
В клинической практике реобазу и хронаксию определяют с помощью метода хронаксиметрии для исследования возбудимости нервных стволов.
4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, Т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя возрастает очень медленно, возбуждения не будет. Это явление называется аккомодацией.
Физиологическая лабильность (подвижность) – это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем выше ее лабильность. Определение лабильности предложено Н.Е. Введенским. Наибольшая лабильность у нервов, наименьшая у сердечной мышцы.
5. Мембрана сост. Из 2 слоев фосфолипидов,по обе стороны-белки. Толщина- 8-10 нм. Мембрана не сплошная, в мембране имеются поры диаметром 0,5-0,7 нм. Через эти поры поступают в-ва и ионы. Поры обладают 2 свойствами: 1- избирательностью (опред.в-во),2- электропровозбудимостью (означ,что проницаемость пор зависит от величины МП) Поры имеют ворота: в условиях покоя А-активационная створка закрыта,но готовы в любую минуту открыться I-инактивационные ворота всегда открыты. В покое поры пропускают только ионы К+,которые имеют все клетки нашего организма,все органоидов клетки. К+ находится в клетке в виде солей. К+ за счет разности концентрации выходит за мембрану, а анионы остаются внутри мембраны и удерживают К+ на мембране. Выход ионов К+ продолжается,пока сила диффузии не приравнивается силам электростатического взаимодействия ( сила= 90mV).
Ходжский (1955) открыл активный механизм,который поддерживает разность концентр.ионов – натриевый и калиевый насос, нах-ся в мембране. Отвечает за обратное поступление ионов К+ из тканевой жидкости в клетку и выделение из нее Na+ против градиента их концентрации в этих средах. Он внутри клетки захватывает 3К+ и выносит на мембрану,затем захватывает 2К+ и переносит их в клетку. Отвечает за обратное
Коэффициенты переноса были выведены Ходжкиным и Хаксли.
Проводимость для калия GK на единицу площади [S/cm²]GK=GKmaxn4 ,где n- фракция К+ каналов в открытом состоянии; GNa=GNa(max)m3h , где m- фракция Na+ каналов в открытом состоянии, h- фракция Na+ каналов в не-инактивированном состоянии
6.Мембранный потенциал - это разность потенциалов между внешней и внутренней сторонами мембраны в условиях, когда клетка не возбуждена. Цитоплазма клетки заряжена отрицательно по отношению к внеклеточной жидкости неравномерным распределением анионов и катионов по обе стороны мембраны. Разность потенциалов (напряжение) для различных клеток имеет значения от -50 до -200 мВ (минус означает, что внутри клетка более отр. заряженная, чем снаружи). Образования и поддержания мембранного потенциала обеспечивают различные типы ионных насосов (в частности натрий-калиевый насос) и ионных каналов (калиевые, натриевые, хлорные ионные каналы). Причина возникновения мембранного потенциала- разность концентрации ионов внешне и внутри клетки. Разность потенциалов рассчитывается уравнением Нернста:
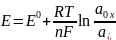 E —
электродный потенциал, E0 —
стандартный электродный потенциал,
измеряется в вольтах;
E —
электродный потенциал, E0 —
стандартный электродный потенциал,
измеряется в вольтах;
R — универсальная газовая постоянная, равная 8.31 Дж/(моль·K);
T — абсолютная температура;
F — постоянная Фарадея, равная 96485,35 Кл·моль−1;
n — число мольэлектронов, участвующих в процессе;
aoxи aRed — активности соответственно окисленной и восстановленной форм вещества, участвующего в полуреакции.
условие- полупроницаемая мембрана одни ионы могут через нее проникать, другие - нет. Оба эти явления зависят от наличия в мембране специальных белков: концентрационные градиенты создают ионные насосы, а проницаемость мембраны для ионов обеспечивают ионные каналы. Важнейшую роль в формировании мембранного потенциала играют ионы K+, Na+ и Cl-. Концентрации этих ионов отличаются по две стороны мембраны. Концентрационные градиенты катионов калия и натрия - это химическая форма потенциальной энергии. В преобразовании энергии в электрическую, участвуют ионные каналы - поры, которые формируются скоплениями специальных трансмембранных белков. Когда ионы диффундируют сквозь канал, они переносят единицу эл. заряда. Любое суммарное движение положительных или отрицательных ионов через мембрану будет создавать напряжение, или разность потенциалов по обе стороны мембраны. Различают пассивный и активный Т. в. через мембраны. В первом случае Т. в. происходит самопроизвольно, при этом молекулы и ионы переносятся в область с более низким электрохимич. потенциалом. Перенос молекул (ионов) в обратном направлении (активный транспорт) возможен только при одновременной затрате энергии, источником к-рой может служить гидролиз АТФ или окислит.-восстановит. реакции в цепях переноса электронов, и осуществляется спец. мол. системами — ионными насосами. Следствием такого активного Т. в., наз. первичным, является неравновесное распределение ионов Н+, Са+, Na+, K+ внутри клетки и между клеткой и окружающей средой; оно, в свою очередь, обеспечивает работу систем сопряжённого, или вторичного активного, Т. в. через мембраны.
7.Потенциал действия, возникающего на мембране электровозбудимых клеток в ответ на раздражение электрическим полем, химическим или другим стимулом. При этом мембрана возбудимой клетки способна увеличивать свою проницаемость к ионам натрия, калия, кальция. Генерация потенциала действия имеет двухфазную циклическую природу. В первой фазе возбуждения происходит реверсия (изменение знака) электрического заряда на мембране — потенциал сдвигается от обычно имеющегося в покое уровня порядка -50 — -90 мВ, до +20 — +40 мВ. Во второй фазе происходит восстановление исходного состояния мембраны, то есть и заряд, и потенциал на мембране быстро возвращаются к исходным значениям, характерным для состояния покоя клетки. Если принять уровень возбудимости в условиях физиологического покоя за норму, то в ходе развития одиночного цикла возбуждения можно наблюдать ее циклические колебания. Периоду статической поляризации соответствует исходная, фоновая возбудимость.
В период развития начальной деполяризации(локальный ответ) на очень короткое время возбудимость незначительно повышается по сравнению с исходной (фаза экзальтации). Во время развития полной деполяризации и инверсии заряда возбудимость падает до нуля. Время, в течение которого отсутствует возбудимость, называется периодом абсолютной рефрактерности: ни один, даже очень сильный раздражитель не может дополнительно вызвать возбуждение ткани. В фазе восстановления мембранного потенциала, т.е. с началом быстрой реполяризации, возбудимость начинает восстанавливаться, но она еще ниже исходного уровня. Время восстановления ее от нуля до исходной величины называется периодом относительной рефрактерности: ткань может ответить возбуждением, но только на сильные, надпороговые раздражения.Вслед за периодом относительной рефрактерности, т.е. с началом фазы медленной реполяризации, наступает короткий период супернормальной— повышенной (по сравнению с исходной) возбудимости.
Заключительный этап одиночного цикла возбуждения — повторное снижение возбудимости ниже исходного уровня (но не до нуля), называемое периодом субнормальной возбудимости, совпадает с развитием гиперполяризации мембраны.
После завершения указанных процессов возбудимость восстанавливается, и клетка готова к осуществлению следующего цикла. Фаза деполяризации- Na+ входит в клетку , фаза реполяризации- К+ выходит из клетки.
8.Возбудимость клетки во время ее возбуждения быстро и сильно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и так же, как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов.
1.Кратковременное повышение возбудимости в начале развития ПД, когда уже возникла некоторая деполяризация клеточной мембраны. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла Екр, при которой возникает ПД.
2.Абсолютная рефрактерная фаза (совпадает с фазой ПД)— это полная невозбудимость клетки (возбудимость равна нулю), она соответствует пику ПД и продолжается 1—2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефрактерная фаза. Клетка в этот период времени на раздражения любой силы не отвечает. Невозбудимость клетки в фазах деполяризации и восходящей части инверсии объясняется тем, что потенциалзависимые т-ворота Na-каналов уже открыты и Na+ быстро поступает в клетку по всем открытым каналам. Те ворота Na-каналов, которые еще не успели открыться, открываются под влиянием деполяризации — умен. мембранного потенциала. Поэтому дополнительное раздражение клетки относительно движения Na+ в клетку ничего изменить не может. Именно поэтому ПД либо совсем не возникает при раздражении, если оно мало; либо является максимальным, если действует раздражение достаточной силы (пороговой или сверхпороговой).
3.Относительная рефрактерная фаза (совпадает с фазой быстрой реполяризации)— это период восстановления возбудимостиклетки, когда сильное раздражение может вызвать новое возбуждение ( Б-3). Относительная рефрактерная фаза соответствует конечной части фазы реполяризации (начиная от Екр ± 10 мВ) и следовой гиперполяризации кл. мембраны, если она имеется. Пониженная возбудимость является следствием все еще повышенной проницаемости для К+ и избыточного выхода его из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, т.к. выход К+ из клетки препятствует ее деполяризации. Кроме того, в период следовой гиперполяризации мембранный потенциал больше и, естественно, дальше отстоит от критического уровня деполяризации. Если реполяризация в конце пика ПД замедляется, то относительная рефрактерная фаза включает и период замедления реполяризации, и период гиперполяризации, т.е. продолжается до возвращения мембранного потенциала к исходному уровню после гиперполяризации. Продолжительность относительной рефрактерной фазы вариабельна, у нервных волокон она невелика и составляет несколько мс.
4.Фаза экзальтации — это период повышенной возбудимости. Он соответствует следовой деполяризации. В некоторых клетках, например в нейронах ЦНС, возможна частичная деполяризация клеточной мембраны вслед за гиперполяризацией. Очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Na+. Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность— это скорость протекания одного цикла возбуждения, т.е. ПД. Как видно из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. При этом особое значение имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани.
9. Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон. Каждое волокно - это клетка цилиндрической формы диаметром 10-100 мкм и длиной от 5 до 400 мкм. Оно имеет клеточную мембрану - сарколемму. В саркоплазме находится несколько ядер, митохондрий образования саркоплазматического ретикулума (СР) и сократительные элементы - миофибрилы. Саркоплазматический ретикулум имеет своеобразное строение. Он состоит из системы поперечных, продольных трубочек и цистерн. Поперечные трубочки это впячивания саркоплазмы внутрь клетки. К ним примыкают продольные трубочки с цистернами. Благодаря этому, потенциал действия может распространяться от сарколеммы на систему саркоплазматического ретикулума. В мышечном волокне содержится более 1000 миофибрилл, расположенных вдоль него. Каждая миофибрилла состоит из 2500 протофибрилл или миофиламентов. Это нити сократительных белков актина и миозина. Миозиновые протофибриллы толстые, актиновые - тонкие. На миозиновых нитях расположены отходящие под углом поперечные отростки с головками. У скелетного мышечного волокна при световой микроскопии видна поперечная исчерченность, т.е. чередование светлых и темных полос. Темные полосы называют А-дисками или анизотропией, светлые 1-дисками (изотропными). В А-дисках сосредоточены нити миозина, обладающие анизотропией и поэтому имеющие темный цвет. 1-диски образованы нитями актина. В центре 1-дисков видна тонкая Z-пластинка. К ней прикрепляются актиновые протофибриллы. Участок миофибрилы между двумя 2-пластинками называется саркомером. Это структурный элемент миофибрилл. В покое толстые миозиновые нити лишь на небольшое расстояние входят в промежутки между актиновыми. Поэтому в средней части А-диска имеется более светлая Н-зона, где нет актиновых нитей. При электронной микроскопии в ее центре видна очень тонкая М-лнния. Она образована цепями опорных белков, к которым крепятся миозиновые протофибриллы.
10. 1 Фаза- электрохимическая стадия
Адекватным раздражителем за счет нервного импульса мышца в месте контактп-синапс выделяется медиатор АХ. Под влиянием АХ повышает проницаемость мембран мышечного волокна для Na и К+. И когда МП мыш.волокна достигает критического уровня деполяризации и возникаеит ПД, этот ПД распространяется до мышечного волокна. ПД вдоль мембраны суммируется, проникает внутрь волокна. Под влиянием ПД из саркоплазмотического ретикулума в саркоплазму выступает Ca.
фаза- сокращение. Кальций => саркоплазма, активирует миозин- они приобретает АТФфазу и милозин расщепляет АТФ. На актине имеются активные центры, которые закрыты белками ( тропомин и тропомиозин). Иона Cа в саркоплазме соединяются с тропонином. Са + тропонин= компекс, и взаимодействуют с тропомиозином и активные центры актина взаимодействуют с мостиками на миозине, и нити актина скользят по нитям миозина. Артомиозин обладает большой активностью.
фаза Расслабления Когда ПД исчезает в раб. вступают Са насос саркоплазмотической ретикулы. И убирает Са из саркоплазмы. Актин отходит от миозина и за счет упругости и эластичности мышца расслабляется
11. Источником энергии для сокращения и расслабления служит АТФ. На головках миозина есть каталитические центры. расщепляющие АТФ до АДФ и неорганического фосфата.. Т.е. миозин является одновременно ферментом АТФ-азой ПД Активность миозина как АТФ-фазы значительно возрастает при его взаимодействии с актином. При каждом цикле взаимодействия актина с головкой миозином расщепляется 1 молекула АТФ. Следовательно, чем больше мостиков переходят в активное состояние, тем больше расщепляется АТФ, тем сильнее сокращение. Для стимуляции АТФ-азной активности миозина требуются ионы кальция, выделяющиеся из СР. которые способствуют освобождению активных центров актина от тропомиозина. Однако запасы АТФ в клетке ограничены. Поэтому для восполнения запасов АТФ происходит его восстановление - ресинтез. Он осуществляется анаэробным и аэробным путем. Процесс анаэробного ресинтеза осуществляется фосфагенной и гликолитической системами. Первая использует для восстановления АТФ запасы креатинфосфата. Он расщепляется на креатин и фосфат, который с помощью ферментов переносится на АДФ (АДФ-Ф=АТФ). Фосфагенная система ресинтеза обеспечивает наибольшую мощность сокращения, но в связи с малым количеством креатинфосфата в клетке, она функционирует лишь 5-6 секунд сокращения. Гликолитическая система использует для ресинтеза АТФ анаэробное расщепление глюкозы (гликогена) до молочной кислоты. Каждая молекула глюкозы обеспечивает восстановление трех молекул АТФ. Энергетические возможности этой системы выше, чем фосфагенной, но и она может служить источником энергии сокращения лишь 0.5 - 2 мин. При этом работа гликолитической системы сопровождается накоплением в мышцах молочной кислоты ;1 снижением содержания кислорода. При продолжительной работе, с усилением кровообращения ресинтез АТФ начинает осуществляться с помощью окислительного фосфолирирования, т.е. аэробным путем. Энергетические возможности окислительной системы значительно больше остальных. Процесс происходит за счет окисления углеводов и жиров. При интенсивной работе в основном окисляются углеводы, при умеренной жиры. Для расслабления также нужна энергия АТФ. После смерти содержание АТФ в клетках быстро снижается и когда становится ниже критического, поперечные мостики миозина не могут отсоединиться от актиновых нитей до ферментативного аутолиза этих белков). Возникает трупное окоченение, АТФ необходима для расслабления потом, что обеспечивает работу Са-насоса
12.При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода:
1.Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мсек. Во время латентного периода генерируется и распространяется ПД, происходит высвобождения кальция из СР, взаимодействие актина с миозином и т.д.
2.Период укорочения. В зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек.,
3.Период расслабления. Его длительность несколько больше, чем укорочения.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются.
13. Суммация - это сложение 2-х последовательных сокращений при нанесении на нее 2-х пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефрактерного периода.
Различают 2 вида суммации: полную и неполную суммацию.
*Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда он уже начала расслабляться.
*Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения. Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить. Например нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности.
14. Тетанус мышцы. Тетанус- это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация. Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений и Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например тремор рук при алкогольной интоксикации и болезни Паркинсона.
15. Утомление - это временное снижение работоспособности мыши в результате работы. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается (рис). Чем выше частота, сила раздражения, величина нагрузки тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, .но/снижается амплитуда (рис.) Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях, полного расслабления не наступает, развивается контрактура. Это состояние непроизвольного длительного сокращения мышцы. Работа утомление мышц исследуются с помощью эргографии. В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления. 1)Теория Шиффа: утомление является следствием истощения энергетических запасов, а мышце.2). Теория Пфлюгера: утомление обусловлено накоплением в мышце продуктов обмена. 3). Теория Ферворна: утомление объясняется недостатком кислорода в мышце. Действительно эти факторы способствуют утомлению в экспериментах на изолированных мышцах. В них нарушается ресинтез ЛТФ. накапливается молочная и пировиноградная кислоты, недостаточно содержание кислорода. Однако в организме интенсивно работающие мышцы, получают необходимый кислород, питательные вещества, освобождаются от метаболитов за счет усиления общего и регионального кровообращения. Поэтому были предложены другие теории утомления. В частности, определенную роль в утомлении принадлежит нервно-мышечным синапсам. Утомление в синапсе развивается из-за истощения запасов нейромедиатора. Однако главная роль, в утомлении двигательного аппарата принадлежит моторным центрам ЦНС. В прошлом веке И.М.Сеченов установил, что если наступает утомление мышц одной руки, то их работоспособность восстанавливается быстрее при работе другой рукой или ногами. Он считал, что это связано с переключением процессов возбуждения с одних двигательных центров на другие. Отдых с включением других мышечных групп он назвал активным. В настоящее время установлено, что двигательное утомление связано с торможением соответствующих нервных центров, в результате метаболических процессов в нейронах, ухудшением синтеза нейромедиаторов и угнетением синаптической передачи.
16. Гладкие мышцы имеются в стенках большинства органов пищеварения, сосудов, выводных протоков различных желёз мочевыводящей системы. Они являются непроизвольными и обеспечивают перистальтику органов пищеварения и мочевыводящей системы, поддержание тонуса сосудов. В отличие от скелетных, гладкие мышцы поперечности клетками чаще веретенообразной формы и небольших размеров, не имеющими поперечной почёрченности. Последнее связано с тем, что сократительный аппарат не обладает упорядоченным строением. Миофибриллы состоят из тонких нитей актина, которые идут в различных направлениях и прикрепляющихся к разным участкам сарколеммы. Миозиновые протофибриллы расположен рядом с актиновыми. Элементы саркоплазматического ретикулума не образуют систему трубочек. Отдельные мышечные клетки соединяются между собой контактами с низким электрическим сопротивлением - нексусами, что обеспечивает распространение возбуждения по всей гладкомышечной структуре. Возбудимость и проводимость гладких мышц ниже, чем скелетных. Мембранный потенциал(МП) составляет 40-60 мВ, так как мембрана ГМК имеет относительно высокую проницаемость для ионов натрия. Причем у многих гладких мышц МП не постоянен. Он периодически уменьшается и вновь возвращается к исходному уровню. Такие колебания называют медленными волнами (МВ). Когда вершина медленные волны, достигает критического уровня деполяризации, на ней начинают генерироваться потенциалы действия, сопровождающиеся сокращениями. МВ и ПД проводятся по гладким мышцам со скоростью всего от 5 до 50 см/сек. Такие гладкие мышцы называют спонтанно активными, т.е. они обладают автоматией. Например, за счет такой активности происходит перистальтика кишечника. Водители ритма кишечной перистальтики расположены в начальных отделах соответствующих кишок.Генерация ПД в ГМК обусловлена входом в них ионов кальция. Механизмы электромеханического сопряжения также отличаются. Сокращение развивается за счет кальция, входящего в клетку во время ПД, Опосредует связь кальция с укорочением миофибрилл важнейший клеточный белок - кальмодулин.Кривая сокращения также отличается. Латентный период, период укорочения, а особенно расслабления значительно продолжительнее, чем у скелетных мышц. Сокращение длится несколько секунд. Гладким мышцам, в отличие от скелетных свойственно явление пластического тонуса. Это способность длительное время находится в состоянии сокращения без значительных энергозатрат и утомления. Благодаря этому свойству поддерживается форма внутренних органов и тонус сосудов. Кроме того, гладкомышечные клетки сами являются рецепторами растяжения. При их натяжении начинают генерироваться ПД, что приводит к сокращению ГМК. Это явление называется: миогенным механизмом регуляции сократительной активности.
17. Взаимодействие нейронов между собой (и с эффекторными органами) происходит через специальные образования - синапсы (греч. - контакт). Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов в нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разнообразных реакциях организма. Особенно много синапсов в высших отделах нервной системы и именно у нейронов с наиболее сложными функциями. В структуре синапса различают три элемента: 1)пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона; 2)синаптическую щель между нейронами; 3)постсинаптическую мембрану - утолщение прилегающей поверхности следующего нейрона. В аксоплазме пресинаптического окончания происходит синтез и накопление медиаторов — веществ, с помощью которых происходит передача импульсов с пресинаптического окончания на постсинаптическую мембрану. Под влиянием нервных импульсов медиаторы высвобождаются из мест накопления (везикул), через пресинаптическую мембрану (часть пресинаптического нервного окончания) поступают в синаптическую щель и воздействуют на чувствительные участки постсинаптической мембраны (рецепторы). Воздействие на рецепторы приводит к соответствующему изменению деятельности клеток, на мембранах которых находятся эти рецепторы. Медиатор может разрушаться в синаптической щели или подвергаться обратному захвату везикулами и вновь использоваться для проведения возбуждения.
Все синапсы можно классифицировать: 1) по их местоположению - центральные (головной и спинной мозг) и периферические; 2) по принадлежности к соответствующим клеткам - нейро-нейрональные, нервно-мышечные, нейро-железистые (нейросекреторные); 3) по месту контакта в нейро-нерональных синапсах - аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и др.; 4) по расположению относительно друг друга (Г.Шеперд) - последовательные синапсы, реципрокные синапсы, синаптические гломерулы (различным способом соединенные через синапсы клетки); 5) по развитию в онтогенезе - стабильные (например, синапсы дуг безусловных рефлексов) и динамические (появляются в процессе индивидуального развития); 6) по знаку их действия - возбуждающие и тормозящие. 7) по способу передачи сигнала - электрические (в которых сигналы передаются электрическим током) и химические (в которых передачиком или посредником является то или иное физиологически активное вещество). Существуют и смешанные - элетрохимические синапсы. 8) химические синапсы классифицируются - по форме контакта: терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона), - по природе медиатора: холинергические (медиатор -ацетилхолин), адренергические (норадреналин), дофаминнергические (дофамин) ГАМК-ергические (гамма-аминомасляная кислота), глицинергические, глутаматергичес- кие, аспартатергические, пуринергические (медиатор -АТФ), - по скорости передачи возбуждения (сигнала): быстрые возбужда-ющие (в передаче принимают участие классические медиаторы потенциал сохраняется короткий промежуток времеми) и медленные (локализованы в спинном мозге, относятся к пептидным синапсам, постсинаптический потенциалы после ритмической стимуляции сохраняются в течение нескольких минут)
18. Передача возбуждения в химическом синапсе – сложный физиологический процесс, протекающий в несколько этапов. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал.
Синтез медиатора
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение синаптических пузырьков в синаптическую щель
Взаимодействие медиатора с рецепторами постсинаптической мембраны
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -каналов. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+ . В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП)
Инактивация медиатора
Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание.
Генерация ПД
В нейро-мышечном синапсе амплитуда единичного ВПСП достаточно велика. Поэтому для генерации ПД в мышечной клетке достаточно прихода одного нервного импульса. Генерация ПД в мышечной клетке происходит в области, окружающей постсинаптическую мембрану.
В нейро-нейрональном синапсе амплитуда ВПСП значительно меньше и недостаточна для того, чтобы деполяризовать мембрану нейрона до КУД. Поэтому для генерации ПД в нервной клетке требуется возникновение нескольких ВПСП. ВПСП, образовавшиеся в результате срабатывания разных синапсов, электротонически распространяются по мембране клетки, суммируются и генерируют образование ПД в области аксонного холмика. Мембрана нейрона в области аксонного холмика обладает низким электрическим сопротивлением и имеет большое количество потенциалчувствительных Na+ -каналов.
19.Нейрон – основная структурная и функциональная единица центральной нервной системы.
Это высокоспециализированные - клетки организма, чрезвычайно различающиеся по своему строению и функциям. В ЦНС нет двух одинаковых нейронов. Мозг человека содержит 25 млрд. нейронов. В общем .плане, все нейроны имеют тело - сому и отростки - дендриты и аксоны. Точной классификации нейронов нет. Но их условно разделяют по структуре и функциям на следующие группы:
1. По форме тела а. Многоугольные б. Пирамидные в. Круглые г. Овальные 2. по количеству и характеру отростков:
* Униполярные - имеющие один отростокб
*Псевдоуниполярные - от тела отходит один отросток, который затем делится на 2 ветви.с.
*Биполярные - 2 отростка, один дендритоподобный, другой аксон. * Мультиполярные - имеют 1 аксон и много дендритов.
3. По медиатору, выделяемому нейроном в синапсе: а). Холинергические б). Адренергические в). Серотонинергические г). Пептидергические и т.д.
4. По функциям: а). Афферентные или чувствительные. Служат для восприятия сигналов из внешней и внутренней среды и передачив ЦНС.
б.) Вставочные или интернейроны, промежуточные. Обеспечивают переработку, хранение и передачу информацияк Афферентным нейронам. Их в ЦНС большинство.
в) Эфферентные или двигательные. Формируют управляющие сигналы, и передают их к периферическим нейронами исполнительным органам.
5. По физиологической роли: а.) Возбуждающие б.) Тормозные
Сома нейронов покрыта многослойной мембраной, обеспечивающей проведение ПД к начальному сегменту аксона -аксонному холмику. В соме расположено ядро, аппарат Гольджи, митохондрни, рибосомы. В рибосомах синтезируются:Тигроид, содержащий РНК и необходимый для синтеза белков. Особую роль играют микротрубочки и тонкие нити -нейрофиламенты. Они имеются в соме и отростках. Обеспечивают транспорт веществ от сомы по отросткам и обратно. Кроме того. за счет нейрофиламентов происходит движение отростков. На дендритах имеются выступы для синапсов - шипики, через которые в нейрон поступает информация. По аксонам сигнал идет к другим нейронам плии волнительным органам.Основными функциями нейронов является их 1.способность к возбуждению. Возбуждение может возникать как в результате синаптических влияний на нейрон других нервных клеток, так и за счет эндогенных цитоплазматических процессов. Внешним выражением возбуждения нейрона является колебание электрического потенциала на его мембране. В невозбужденном нейроне регистрируется мембранный потенциал, или потенциал покоя, около —70 мВ.2. синтез БАВ3. 2. воспроизведение информации3. хранение и интеграция информации в пресинаптических окончаниях. 4. в аксоне: аксонный транспорт, генерация электрических импульсов, выделение медиатора. Каждый нейрон синтезирует в своем теле и затем выделяет во всех своих синапсах один и тот же медиатор, поэтому нейроны и ацетилхолиновой передачей возбуждения называются холинергическими, с адреналиновой – адренергическими.. Норадренергические нейроны обнаружены в составе среднего мозга, моста и продолговатого мозга. В состав дорсального и медиального ядер продолговатого мозга, моста и среднего мозга входят серотонические нейроны Наличие кальция в нервной клетке имеет отношение к перераспределению ионов Na+ и К+ в клетке, синтезу и секреции медиаторов, синтезу белка и РНК, аксоплазматическому транспорту. При синаптической активации постсинаптических мембран из них выделяются простагландины, которые изменяют энергетический метаболизм нейронов, участвуют в регуляции возбудимости клетки, секреции медиаторов и гормонов.
20. Нейроглия — структурный компонент нервной ткани, окружающий нервную клетку и не обладающий способностью к проведению нервных импульсов. Нейроглия выполняет ряд функций, которые обеспечивают нормальную деятельность нейронов. В нейроглии различают макроглию, микроглию и олигодендроглию. В состав макроглии входят астроглия, олигодендроглия и эпендима. Астроглия построена из звездчатых клеток — астроцитов, выполняющих трофическую и опорную функции. Их отростки формируют сеть, в петлях которой лежат нейроны. Концевые отростки астроцитов подходят к кровеносным сосудам, изолируя их от нейронов. Эпендимная глия выстилает центральный канал спинного мозга и полости мозговых желудочков. Олигодендроглия построена из клеток олигодендроцитов, имеющих слабо ветвящиеся отростки. Олигодендроциты секретируют миелин, участвуют в трофике нейронов, имеют отношение к водному обмену нервной ткани. Микроглия представлена микроглиоцитами — клетками с короткими отростками, на которых имеются мелкие выросты. Клетки микроглии выполняют фагоцитарную функцию.
Существуют следующие методы исследования функций ЦНС:
1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом.
2. Метод экстирпации (удаления) или разрушения участков мозга.
3. Метод раздражения различных отделов и центров мозга.
4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием.
5. Электрофизиологические методы:
а. электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г. Бергером.
б. регистрация биопотенциалов различных нервных центров; используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро.
в. метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков;
6. метод внутримозгового введения веществ с помощью микроинофореза;
7. хронорефлексометрия – определение времени рефлексов.
21. Простейшим нервным центром является нервная цепь, состоящая из трех последовательно соединенных нейронов. Нейроны сложных нервных центров имеют многочисленные связи между собой, образуя нервные сети трех типов:
1. Иерархические. Если возбуждение распространяется на все большее количество нейронов, то такое явление называется дивергенцией (рис). Если же наоборот, от нескольким нейронов пути идут к меньшему количеству, такой механизм называется конвергенцией (рис). Например, к одному мотонейрону могут подходить нервные окончания от нескольких афферентных нейронов. В таких сетях вышележащие нейроны управляют ниже лежащими.
2. Локальные сети. Содержат нейроны с короткими аксонами. Они обеспечивают связь нейронов очного уровня ЦНС II кратковременное сохранение информации на этом уровне. Примером их является кольцевая цепь (рис). По таким цепям возбуждение циркулирует определенное время. Такая циркуляция называется реверберацией возбуждения (мех. кратковременной памяти).
3. Дивергентные сети с одним входом. В них один нейрон, т.е. вход образует большое количество связей с нейронами многих центров.
В связи с наличием многочисленных связей между нейронами сети в них может возникать иррадиация возбуждения. Это его распространение на все нейроны. В результате иррадиации возбуждение может переходить на другие нервные центры и даже охватывать всю нервную систему.
В нервных сетях большое количество вставочных нейронов, ряд из которых является тормозными. Поэтому в них может возникать несколько типов тормозных процессов:
1) Реципрокное торможение. В этом случае, сигналы идущие от афферентных нейронов, возбуждают одни нейроны. но одновременно, через вставочные тормозные нейроны, тормозят другие. Такое торможение называется также сопряженным (рис).
Возвратное торможение. При этом, возбуждение идет от нейрона по аксону к другой клетке. Но одновременно по коллатералям (ветвям) к тормозному нейрону, который образует синапс на теле этого же нейрона. Частный случай такого торможения - торможение Реншоу. При возбуждении мотонейронов спинного мозга, нервные импульсы по их аксонам идут к мышечным волокнам, но одновременно они распространяются по коллатералям этого аксона к клокам Реншоу. Аксоны клеток Реншоу образуют тормозные синапсы на телах этих же мотонейронов. В результате, чем сильнее возбуждается мотонейрон, тем более сильное тормозящее влияние на него оказывает тормозной нейрон Реншоу (рис). Такая связь в ЦНС называется обратной отрицательной.
Латеральное торможение. Это процесс, при котором возбуждение одной нейронной цепи приводит к торможению параллельной с такими же функциями. Осуществляется через вставочные нейроны.
Свойства нервных центров :
Рассматриваемые ниже свойства нервных центров связаны с некоторыми особенностями распространения возбуждения в ЦНС, особыми свойствами химических синапсов и свойствами мембран нервных клеток. Основными свойствами нервных центров являются следующие.
А. Инерционность — сравнительно медленное возникновение возбуждения всего комплекса нейронов центра при поступлении к нему импульсов и медленное исчезновение возбуждения нейронов центра после прекращения входной импульсации. Инерционность центров связана с суммацией возбуждения и последействием.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры, процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Явление суммации возбуждения в ЦНС открыл И.М.Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождаются ответной реакцией: лягушка совершает прыжок. Различают временную (последовательную) и пространственную суммацию. Временная суммация обусловлена тем, что ВПСП (возбуждающие постсинаптические материалы) от предыдущего импульса еще продолжается, когда приходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией. Она играет важную физиологическую роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в нейронных объединениях нервных центров. Пространственная суммация , раздельная стимуляция каждого из двух аксонов вызывает под-пороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД — процесс, который не может быть обеспечен одиночным ВПСП. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция. 3. Последействие — это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Последействие является результатом ряда причин.
• Длительное существование ВПСП, если ВПСП является полисинаптическим и высокоамплитудным. В этом случае при одном ВПСП возникает несколько ПД.
• Многократные появления следовой деполяризации, что свойственно нейронам ЦНС. Если следовая деполяризация достигает Екр, то возникает ПД.
• Циркуляция возбуждения по замкнутым нейронным цепям (см. рис. 7.4). Первые две причины действуют недолго: десятки — сотни миллисекунд, третья причина (циркуляция возбуждения) может продолжаться минуты и даже часы. Таким образом, особенность распространения возбуждения (его циркуляция) обеспечивает другое явление в ЦНС — последействие. Последействие играет важнейшую роль в процессах обучения, в том числе кратковременной памяти.
4. Фоновая активность нервных центров
(тонус) объясняется: 1) спонтанной активностью нейронов ЦНС; 2) гуморальными влияниями биологически активных веществ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и влияющих на возбудимость нейронов; 3) афферентной им-пульсацией от различных рефлексогенных зон; 4) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах; 5) циркуляцией возбуждения в ЦНС. Значение фоновой активности нервных центров заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов. Этот уровень может увеличиваться или уменьшаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора.
5. Трансформация ритма возбуждения — изменение числа импульсов, возникающих в нейронах центра на выходе относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна в сторону как увеличения, так и уменьшения. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют иррадиация процесса возбуждения (см. раздел 7.6) и последействие. Уменьшение числа импульсов в нервном центре объясняется снижением его возбудимости за счет процессов пре- и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных влияний, когда уже все нейроны центра или нейронного пула возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов и нервных импульсов на выходе.
6. Большая чувствительность ЦНС к изменениям внутренней среды, например к изменению содержания глюкозы в крови, газового состава крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы могут возникнуть судороги. Тяжелые последствия для ЦНС вызывает недостаток кислорода в крови. Прекращение кровотока всего лишь на 10 с приводит к очевидным нарушениям функций мозга, человек теряет сознание. Прекращение кровотока на 8—12 мин вызывает необратимые нарушения деятельности мозга: погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям.
7. Утомляемость нервных центров продемонстрировал Н.Е.Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмическое сокращение мышцы, приводящее к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги (рис. 7.6), при этом развивается синоптическая депрессия — ослабление реакции центра на раздражения (афферентные импульсы), выражающееся в снижении постси-наптических потенциалов во время длительного раздражения или после него. Она объясняется расходованием медиатора, снижением чувствительности постсинаптической мембраны к медиатору вследствие накопления метаболитов, закисления среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
8. Пластичность нервных центров — способность нервных элементов к перестройке функциональных свойств. Основные проявления этого свойства следующие.
9. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы,. выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
22. Явление центрального торможения обнаружено И.М. Сеченовым в 1362 году. Он обнаружил, что если на поперечный разрез зрительных бугров лягушки наложить кристаллик поваренной соли или подействовать электрическим слабым током, то время рефлекса Тюрка резко удлиняется (рефлекс Тюрка - сгибание лапки при погружении ее у в кислоту). Вскоре были открыты новые факты, демонстрирующие явления торможения в ЦНС. Гольц показал, что рефлекс Тюрка затормаживается при сдавливании пинцетом другой лапки, Шеррингтон доказал наличие торможения рефлекторного сокращения разгибателя при осуществлении сгибательного рефлекса. Было доказано, что при этом интенсивность рефлекторного торможения зависит от соотношения силы возбуждающего и тормозящего раздражителей.
Торможение в ЦНС препятствует развитию возбуждения или ослабляет протекающее возбуждение. Примером торможения может быть прекращение рефлекторной реакции, на фоне - действия другого более сильного раздражителя. Первоначально была предложена унитарно-химическая теория торможения. Она основывалась на принципе Дейла: один нейрон - один медиатор. Согласно ей торможение обеспечивается теми же нейронами и синапсами, что и возбуждение. В последующем была доказана правильность бинарно-химической теории. В соответствии с последней, торможение обеспечивается специальными тормозными нейронами, которые являются вставочными. Это клетки Реншоу спинного мозга и нейроны Пуркинье промежуточного. Торможение в ЦНС необходимо для интеграции нейронов в единый нервный центр.
Постсинаптическое торможение.
Наибольшее распространение в ЦНС имеет механизм постсинаптического торможения, которое осуществляется специальными тормозными вставочными нервными клетками (например, клетки Реншоу в спинном мозге или клетки Пуркинье (грушевидные нейроны) в коре мозжечка). Особенность тормозных нервных клеток состоит в том, что в их синапсах имеются медиаторы, вызывающие на постсинаптической мембране нейрона ТПСП (тормозящие постсинаптические потенциалы), т.е. кратковременную гиперполяризацию. Например, для мотонейронов спинного мозга гиперполяризующим медиатором является аминокислота глицин, а для многих нейронов коры большого мозга таким медиатором служит гамма-аминомасляная кислота — ГАМК. Частным случаем постсинаптического является возвратное торможение.
Постсинаптическое. Оно возникает в постсинаптической мембране сомы и дендритов нейронов, т.е. после передающего синапса. На этих участках образуют аксо-дендритные или аксосоматические синапсы специализированные тормозные нейроны (рис). Эти синапсы являются глицинергическими. В результате воздействия, НЛИ на глициновые хеморецепторы постсинаптической мембраны, открываются, ее калиевые и хлорные каналы. Ионы калия и хлора входят в нейрон, развивается ТПСП. Роль ионов хлора в развитии ТПСП: небольшая. В результате возникшей гиперполяризации возбудимость нейрона падает. Проведение нервных, импульсов через него прекращается. Алкалоид стрихнин может связываться с глицериновыми рецепторами постсинаптической мембраны и выключать тормозные синапсы. Это используется для демонстрации роли торможения. После введения стрихнина у животного развиваются судороги всех мышц.
Виды постсинаптического торможения
В основе классификации постсинаптического торможения лежит путь, по которому тормозная клетка вовлекается в ответную реакцию.
Прямое (афферентное, поступательное) торможение - возникает когда тормозная клетка получает импульсы от афферентного нейрона или от вышележащих отделов центральной нервной системы.
Возвратное (эфферентное) торможение - клетки Реншоу получают импульсы по коллатералям аксона эфферентного нейрона. Эфферентный нейрон образует аксон, который иннервирует скелетные мышцы. От этого аксона отходит ответвление, которое образует синапс на клетке Реншоу. Клетка Реншоу тормозит нейрон от которого получает нервный импульс.
Пресинаптическое торможение. Пресинаптическое торможение развивается в пресинаптической части синапса за счет воздействия на его мембрану аксо-аксональных синапсов. В результате как деполяризующего, так и гиперполяризующего воздействия происходит блокирование проведения импульсов возбуждения по пресинаптическим путям к постсинаптической нервной клетке.
Пресинаптическое торможение. В этом случае тормозной нейрон образует синапс на аксоне нейрона, подходящем к передающему синапсу. Т.е. такой синапс является аксо-аксональным (рис). Медиатором этих синапсов служит ГАМК. Под действием ГАМК активируются хлорные каналы постсинаптической мембраны. Но в этом случае ионы хлора начинают выходить из аксона. Это приводит к небольшой локальной, но длительной деполяризации его мембраны.
Значительная часть натриевых каналов мембраны инактивируется, что блокирует проведение нервных импульсов по аксону, а следовательно выделение нейромедиатора в передающем синапсе. Чем ближе тормозной синапс расположен к аксонному холмику, тем сильнее его тормозной эффект. Пресинаптическое торможение наиболее эффективно при обработке информации, так как проведение возбуждения блокируется не во всем нейроне, а только на его одном входе. Другие синапсы, находящиеся на нейроне продолжают функционировать.
Пессимальное торможение. Торможение деятельности нервной клетки может осуществляться и без участия особых тормозных структур. В этом случае оно возникает в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под влиянием слишком частых импульсов (как пессимум в нервно-мышечном препарате). К пессимальному торможению особо склонны промежуточные нейроны спинного мозга, нейроны ретикулярной формации. При стойкой деполяризации в них наступает состояние, подобное катодической депрессии Вериго.
Выделяют первичное и вторичное торможение.
Первичное (механизм центрального торможения):
постсинаптический
пресинаптический
В процессе постсинаптического торможения процесс идет на постсинаптической мембране, возникает гиперполяризация за счет тормозного медиатора глицина. Поэтому блокируется весь нейрон одновременно.
Пресинаптическое торможение осуществляется путем вытормаживания какого-то определенного пути, идущего к данному нейрону.
Выделяют:
1) рецепроктное торможение (Н: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя – открыл Шеррингтон.
2) Возвратное торможение. Н: альфа – мотонейрон посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС – она заканчивается на тормозном нейроне и активирует её. Тормозной нейрон выщывает торможение альфа –мотонейрона, который запустил всю эту цепочку. Т.е. альфа –мотонейрон , активируясь через систему тормозного нейрона сам себя тормозит.
3) существует ряд вариантов возвратного тормажения, в частности латеральное тормажение. Н: активируется фоторецептор, он активирует биполярную клетку и одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглинозной клетке. Этим самым происходит затормаживание информации в соседних участках. Создаются условия для четкого видения предмета.
Вторичное тормажение:
1) пессимальное – связано с истощением количества медиатора, т.е происходит удлинение абсолютной рефрактерной фа0зы (явление стойкой деполяризации). Присуще вставочным нейронам, т.к. они обладают низкой лабильностью.
2) торможение вслед за возбуждением. Связано со следовой гиреполяризацией. Возбудимость пониженная (т.е. клетка не может ответить возбуждением).
Торможение является фактором, способствующим приспособлению организма к окружающим его условиям. Так же торможение ослабляет процессы возбуждения в нервной системе и обеспечивает стабильность её работы. При отсутствии торможения, процессы возбуждения нарастали бы и накапливались, что неминуемо привело бы к разрушению нервной системы и гибели организм.
23. Рефлекторная реакция в большинстве случаев осуществляется не одной, а целой группой рефлекторных дуг и нервных центров. Координация рефлекторной деятельности это такое взаимодействие нервных центров и проходящих по ним нервных импульсов, которое обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов:
1. Временное и пространственное облегчение. Это усиление рефлекторной реакции при действии ряда последовательных раздражителей или одновременном их воздействии на несколько рецептивных полей. Объясняется явлением суммации в нервных центрах.
2. Окклюзия явление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне.
3. Принцип общего конечного пути. Разработан Ч. Шеррингтоном. В основе его лежит явление конвергенции. Согласно этому принципу на одном эфферентном мотонейроне могут образовывать синапсы нескольких афферентных, входящих в несколько рефлекторных дуг. Этот нейрон называется общим конечным путем и участвует в нескольких рефлекторных реакциях. Если взаимодействие этих рефлексов приводит к усилению обшей рефлекторной реакции, такие рефлексы называются союзными. Если же между афферентными сигналами происходит борьба за мотонейрон - конечный путь, то антагонистическими. В результате этой борьбы второстепенные рефлексы ослабляются, а жизненно важным освобождается общий конечный путь.
4. Реципрокное торможение. Обнаружено Ч. Шеррингтоном. Это явление торможения одного Центра в результате возбуждения другого. Т.е. в этом случае тормозится антагонистический центр. Например при возбуждении центров сгибания левой ноги по реципрокному механизму тормозятся центры мышц разгибателей этой же ноги и центры сгибателей правой. В реципрокных взаимоотношениях находятся, центры вдоха и выдоха продолговатого мозга. центры сна и бодрствования и т.д.
5. Принцип доминанты. Открыт А.А. Ухтомским. Доминанта - это преобладающий очаг возбуждения в ЦНС, подчиняющий себе другие НЦ. Доминантный центр обеспечивает комплекс рефлексов, которые необходимы в данный момент для достижения определенной цели. При некоторых условиях возникают питьевая, пищевая, оборонительная, половая и др. доминанты. Свойствами доминантного очага являются повышенная возбудимость, стойкость возбуждения, высокая способность к суммации, инертность. Эти свойства обусловлены явлениями облегчения, иррадиации, с одновременным повышением активности вставочных тормозных нейронов, которые тормозят нейроны других центров.
6. Принцип обратной афферентации. Результаты рефлекторного акта воспринимаются нейронами обратной афферентации и информация от них поступает обратно в нервный центр. Там они сравниваются с параметрами возбуждения и рефлекторная реакция корректируется.
24. Строение. Расположен спинной мозг в позвоночном канале от I шейного позвонка до I — II поясничных, длина около 45 см, толщина около 1 см. Передняя и задняя продольные борозды делят его на две симметричные половинки. В центре проходит спинномозговой канал, в котором находится спинномозговая жидкость. В средней части спинного мозга, около спинномозгового канала расположено серое вещество, на поперечном срезе напоминающее контур бабочки. Серое вещество образовано телами нейронов, в нем различают передние и задние рога. В задних рогах спинного мозга расположены тела вставочных нейронов, в передних — тела двигательных нейронов. В грудном отделе различают еще и боковые рога, в которых расположены нейроны симпатической части автономной нервной системы. Вокруг серого вещества расположено белое вещество, образованное нервными волокнами. Спинной мозг покрыт тремя оболочками: снаружи соединительно-тканная плотная, затем паутинная и под ней сосудистая. От спинного мозга отходят 31 пара смешанных спинномозговых нервов. Каждый нерв начинается двумя корешками, передним (двигательным), в котором находятся отростки двигательных нейронов и вегетативные волокна, и задним (чувствительным), по которому возбуждение передается к спинному мозгу. В задних корешках находятся спинномозговые узлы, скопления тел чувствительных нейронов. Перерезка задних корешков приводит к утрате чувствительности в тех областях, которые иннервируются соответствующими корешками, перерезка передних корешков приводит параличу иннервируемых мышц.
Спинной мозг выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями. Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, передние - двигательными. Эта закономерность называется законом Белла-Мажанди. Корешки каждого сегмента иннервируют 3 метамера тела, но в результате перекрывания каждый метамер иннервируется тремя сегментами. Поэтому при поражении передних корешков одного сегмента, двигательная активность соответствующего метамера лишь ослабляется. Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов внс. Мотонейроны функционально разделяются на альфа- и гамма-мотонейроны. К мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных. т.е. рецепторных мышечных клеток. Тела а-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом, они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает. Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС. Нейроны симпатического отдела вегетативной нервной системы находятся в боковых рогах грудных сегментов, а парасимпатического в крестцовом отделе. Все рефлексы спинного мозга делятся на соматические и вегетативные. Соматические делятся на сухожильные или миотатические и кожные. Сухожильные возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и а-мотонейронов. В результате- сокращение мышц, в первую очередь разгибателей. К сухожильным рефлексам относятся коленный, ахиллов, локтевой, кистевой и др. Например, коленный является простейшим моносинаптическим, так как в его центральной части только один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной. Вегетативные р-сы делятся на симпатические и парасимпатические. Те и другие проявляются, реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эрекции и т.д
По чувствительным нервным волокнам спинной мозг получает сигналы от экстерорецепторов поверхности кожи, проприорецепторов мышц туловища и конечностей и от интерорецепторов внутренних органов. Эти сигналы передаются двигательным нейронам спинного мозга, распространяются по их аксонам и регулируют сократительную деятельность всей мускулатуры тела человека, за исключением мышц шеи и головы, иннервируемых черепными нервами. Двигательные центры спинного мозга обеспечивают также постоянный тонус мышц, т. е. некоторое постоянное их напряжение, не исчезающее даже во время сна. Рефлекторный тонус мышц имеет очень важное значение для движения конечностей и сохранения определенного положения тела (стоя, сидя, лежа, при наклоне и т. д.).В боковых рогах грудного и верхних сегментах поясничного отделов спинного мозга расположены центры симпатического отдела вегетативной нервной системы, иннервирующие сосуды, потовые железы, сердце, пищеварительный тракт и другие внутренние органы, скелетные мышцы, т. е. по существу все органы и ткани организма. В боковых рогах крестцового отдела спинного мозга расположены центры парасимпатического отдела вегетативной нервной системы, иннервирующие органы малого таза и являющиеся центрами мочеиспускания, дефекации и некоторых других функций. Сигналы, поступающие в спинной мозг от рецепторов, передаются по многочисленным проводящим путям, расположенным в задних и боковых канатиках спинного мозга, в центры ствола головного мозга, достигают коры больших полушарий и мозжечка. В свою очередь от расположенных выше отделов центральной нервной системы в спинной мозг приходят импульсы по проводящим путям передних и боковых канатиков. Эти сигналы оказывают возбуждающее или тормозящее действие на вставочные и двигательные нейроны спинного мозга, благодаря чему изменяется деятельность скелетной мускулатуры и внутренних органов. Каждый сегмент спинного мозга иннервирует три поперечных отрезка, или метамера, тела: свой собственный, один выше и один ниже. Таким образом, каждый метамер тела получает чувствительные волокна от трех корешков. Скелетные мышцы тоже снабжаются двигательными волокнами от трех сегментов спинного мозга.
25. Проводниковая функция спинного мозга, состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные, восходящие и нисходящие. Собственные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища. К восходящим путям относятся несколько трактов. Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности. Нисходящие, пути также образованы несколькими трактами. Кортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры а-мотонеронам спинного мозга. За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга, к а-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к а-мотонейронам мышц разгибателей. За счет этих двух путей и регулируется тонус соответствующих мышц при изменениях, положения тела. Все рефлексы спинного мозга делятся на соматические и вегетативные. Соматические делятся на сухожильные или миотатические и кожные. Сухожильные-возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и а-мотонейронов. В результате- сокращение мышц, в первую очередь разгибателей. К сухожильным рефлексам относятся коленный, ахиллов, локтевой, кистевой и др. Например, коленный является простейшим моносинаптическим, так как в его центральной части только один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной. Вегетативные р-сы делятся на симпатические и парасимпатические. Те и другие проявляются, реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эрекции и т.д.Спинальные нц находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится.
26. Продолговатый мозг - продолжение спинного и повторяет его строение: на передней и задней поверхности здесь также залегают борозды. Он состоит из белого вещества (проводящих пучков), где рассеяны скопления серого вещества - ядра, от которых берут начало черепные нервы - с IX по XII пару, в их числе языкогло-точный (IX пара), блуждающий (X пара), иннервирующий органы дыхания, кровообращения, пищеварения и другие системы, подъязычный (XII пара). Вверху продолговатый мозг продолжается в утолщение - варолиев мост, а с боков от него отходят нижние ножки мозжечка. Сверху и с боков почти весь продолговатый мозг прикрыт большими полушариями и мозжечком. В сером веществе продолговатого мозга залегают жизненно важные центры, регулирующие сердечную деятельность, дыхание, глотание, осуществляющие защитные рефлексы (чихание, кашель, рвота, слезоотделение), секрецию слюны, желудочного и поджелудочного сока и др. Повреждение продолговатого мозга может быть причиной смерти вследствие прекращения сердечной деятельности и дыхания. Функционально в нем выделяют 2 отдела: таламус и гипоталамус. В таламусе происходит обработка почти всей информации, идущей от рецепторов к коре. Сам он содержит около 120 ядер. Они делятся на неспецифические и специфические. Неспецифические относятся к переднему отделу ретикулярной формации ствола мозга. Их аксоны нейронов поднимаются к коре и диффузно пронизывают все ее слои. К этим ядрам подходят нервные волокна от нижележащих отделов гипоталамуса, лимбической системы, базальных ядер. При возбуждении неспецифических ядер в коре мозга развивается периодическая электрическая активность в виде веретен, что свидетельствует о переходе ко сну, т.е. они обеспечивают определенный уровень функционального активности коры. Специфические ядра делятся на переключающие или релейные и ассоциативные. Переключающие ядра состоят из нейронов, у которых мало дендритов и длинный аксон. С помощью них происходит переключение сигналов идущих от нижележащих отделов ЦНС на соответствующие соматосенсорные зоны коры. в которых находится представительство определенных рецепторов. Например, в латеральных коленчатых телах переключаются зрительные сигналы на затылочные доли коры. В переключающих ядрах выделяется наиболее важная информация. При нарушении функции этих ядер выключается восприятие соответствующих сигналов. Ассоциативные нейроны имеют большее количество отростков и синапсов. Это позволяет им воспринимать различные по характеру сигналы. Они их получают эти сигналы от переключающих и осуществляют их первичный синтез. От них пути идут к ассоциативным зонам коры, в которых происходит высший синтез, и формируются сложные ощущения. Кроме того, ядра таламуса участвуют в формировании безусловных двигательных рефлексов сосания, жевания, глотания. В таламусе находится подкорковый центр болевой чувствительности, в котором формируется общее ощущение боли, не имеющее определенной локализации и окраски. Верхние бугры четверохолмия являются первичными зрительными центрами. К ним подходят пути от нейронов сетчатки глаза. От них сигналы идут к таламусу, а по нисходящему тектоспинальному пути к мотонейронам спинного мозга. Здесь происходит первичный анализ зрительной информации. Например, определение положения источника света направление его движения. В них также формируются зрительные ориентировочные рефлексы. т.е. поворот головы в сторону источника света. Нижние бугры четверохолмия являются первичными слуховыми центрами. К ним идут сигналы от фонорецепторов уха. а от них к таламусу. От них к мотонейронам также идут пути в составе гектоспинального тракта. В этих буграх осуществляется первичный анализ слуховых сигналов, а за счет связей с мотонейронами формируются ориентировочные рефлексы на звуковые раздражители. Основными функциями продолговатого мозга являются проводниковая, рефлекторная и ассоциативная. Первая осуществляется проводящими путями, проходящими через него. Вторая, нервными центрами. В ромбовидной ямке продолговатого мозга находятся ядра 10, 11, 12 пар черепно-мозговых нервов, а также ретикулярная формация. Рефлекторные функции делятся на соматические и вегетативные. Соматическими являются статистические рефлексы продолговатого мозга, относящиеся к позно-тоническим или рефлексам позы. Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальные тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышаемся тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности. В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный, сосудодвигательный центры и центр регуляции сердечной деятельности. Первый обеспечивает смену фаз дыхания, второй - тонус периферических сосудов, третий регуляцию частоты и силы сердечных сокращений. В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени. Здесь же расположены центры регуляции моторики пищеварительного канала. Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный центр, центры кашля, чихания, смыкания век и слезотечения при раздражении роговицы. Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов - сосания, жевания, глотания. В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности, в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
27. Средний мозг соединяет все отделы головного мозга. Здесь находятся центры тонуса скелетных мышц, первичные центры зрительных и слуховых ориентировочных рефлексов. Эти рефлексы проявляются в движениях глаз, головы в сторону раздражителей. Средний мозг состоит из крыши среднего мозга, ножек мозга и полости среднего мозга. Крыша среднего мозга – самая главная часть в строении его. Именно в крыше располагаются промежуточные центры зрения и слуха. Центры эти располагаются в виде 4-х холмиков: 2 верхние и 2 нижние. Верхние холмики отвечают за зрение, нижние – за слух.
Мост тесно связан со средним мозгом. Осуществляют проводниковую и рефлекторную функции. Проводниковая обеспечивается восходящими и нисходящими путями, идущими через них. Рефлекторная их нервными центрами. Нейроны моста формируют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательную часть ядер тройничного нерва и его чувствительное ядро. Ретикулярная формация является частью всей ретикулярной формации ствола. В ней расположены ядра пневмотаксического центра. Он координирует активность центров вдоха и выдоха продолговатого мозга. В среднем мозге находятся ядра четверохолмия, красное ядро, черная субстанция, ядра глазодвигательного и блокового нервов, ретикулярная формация. Красное ядро расположены в верхней части ножки мозга. К нему идут нервные пути от коры полушарий, подкорковых ядер, мозжечка. От него идет руброспинальный тракт к мотонейронам сгибателей спинного и ретикулярной формации продолговатого мозга. В связи с различным функциональным значением ядра Дейтерса и красного ядра, при перерезке ствола между средним и продолговатым мозгом у животных возникает децеребрационная ригидность. Это резкое повышение тонуса всех мышц разгибателей. Голова животного запрокидывается, спина выгибается, конечности вытягиваются. Механизм Д.Р. заключается в том. что красное ядро, активируя мотонейроны сгибателей, через вставочные тормозные нейроны тормозит мотонейроны разгибателей. Одновременно исключается тормозящее влияние красного ядра на ретикулярную формацию продолговатого мозга, возле ядра Дейтерса. В отсутствии влияния красного ядра преобладает возбуждающее действие ядра Дейтерса на мотонейроны разгибателей.Позные рефлексы продолговатого и выпрямительные среднего мозга, обеспечивают непроизвольное поддержание позы и равновесия тела при неподвижном положении, например стоянии, сидении. Поэтому они относятся к статическим.Статокинетические рефлексы. Это рефлексы, которые служат для сохранения устойчивого положения тела при движении. К ним относятся нистагм головы и глаз, лифтная реакция, рефлекс готовности к прыжку. Нистагм головы и глаз это их медленное бессознательное движение в сторону противоположную вращению, а затем быстрое возвращение в исходную позицию. Нистагм глаз сохраняется некоторое время, и после вращения. Лифтная реакция - это уменьшение тонуса разгибателей конечностей в начале быстрого подъема, которое сменяется его повышением. При быстром опускании, тонус разгибателей меняется противоположным образом. Рефлекс готовности к прыжку проявляется увеличением тонуса разгибателей передних конечностей при опускании животного вниз головой. В результате они вытягиваются. Статокинетические рефлексы как и выпрямительные, обусловлены возбуждением рецепторов вестибулярного аппарата.Ядра глазодвигательного и блокового нервов обеспечивают содружественные движения глаз. Кроме, того первое регулирует ширину зрачка и кривизну хрусталика. Черная субстанция имеет двусторонние связи с подкорковыми ядрами и участвует в координации точных движений пальцев рук, регуляции жевания и глотания. Она может оказывать тормозное влияние на красное ядро.
28.. В мозжечке выделяют три части: парные полушария и соединяющее их образование – червь. Поверхность мозжечка покрыта серым веществом – корой. Кора полушарий и червь пересечены параллельными щелями мозжечка, разделяющими его листки, или извилины. Местами имеются более глубокие борозды, ограничивающие дольки мозжечка, симметричные на обоих полушариях. Среди всех долек выделяют клочок – самую филогенетически старую дольку, соприкасающуюся со средней мозжечковой ножкой; клочок имеет ножку, посредством которой сообщается с червем .С другими отделами головного мозга строение мозжечка соединяют три пары ножек. Нижние идут к продолговатому мозгу, средние – к мосту, верхние переходят в средний мозг. В составе ножек в мозжечок следуют волокна восходящих и нисходящих проводящих путей. Полушария и червь снаружи покрыты серым веществом, а внутри содержат белое. В толще листков полушарий проходят тонкие полоски белого вещества. Поперечный разрез мозжечка, разделенного бороздами, имеет форму дерева, поэтому его называют «древо жизни». Мозжечок получает афферентные нервные импульсы от тактильных рецепторов, рецепторов вестибулярного аппарата, проприорецепторов мышц и сухожилий, а также двигательных зон коры. Эфферентные импульсы от мозжечка идут к красному ядру среднего, ядру Дейтерса продолговатого мозга, к таламусу, а затем моторным зонам коры и подкорковым ядрам. Общей функцией мозжечка является регуляция позы и движений. Эту функцию он осуществляет путем координации активности других двигательных центров:
вестибулярных ядер, красного ядра, пирамидных нейронов коры. Поэтому он выполняет следующие двигательные функции: 1. Регуляцию мышечного тонуса и позы. 2. Коррекцию медленных целенаправленных движений в ходе их выполнения, а также координацию этих движений с рефлексам и положения тела. 3. Контроль за правильным выполнением быстрых движений, осуществляемых корой. В связи с тем, что мозжечок выполняет данные функции, при его удалении у животного развивается комплекс двигательных нарушений, называемый триадой Лючиани. Он включает:1. Агония и дистония - снижение и неправильное распределение тонуса скелетных мышц.2. Астазия - невозможность слитного сокращения мыши. а как следствие, сохранения устойчивого положения тела при стоянии, сидении (покачивание).3. Астения - быстрая утомляемость мыши.4. Атаксия - плохая координация движений при ходьбе. Неустойчивая "пьяная" походка.5. Адиадохокинез - нарушение правильной последовательности быстрых целенаправленных движений.
В клинике умеренные поражения мозжечка проявляются триадой Шарко:
1. Нистагм глаз в состоянии покоя. 2. Тремор конечностей, возникающий при их движениях. 3.Дизартрия - нарушения речи. Орбели установил, что мозжечок влияет и на различные вегетативные функции, эти влияния могут, быть возбуждающими и тормозящими. Например, при раздражении мозжечка увеличивается .или снижается кровяное давление, изменяется частота сердцебиений, дыхание, пищеварение. Мозжечок влияет на обмен веществ. На, эти функции он воздействует через вегетативные нервные центры, координируя их активность с движением. функции внутренних органов изменяются в связи с изменением обменных процессов в них. Поэтому мозжечок оказывает на них адаптационно-трофическое влияние.
29. Ретикулярная формация (РФ) представляет собой сложное скопление нервных клеток, характеризующихся обширно разветвленным дендритным деревом и длинными аксонами, часть из которых имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее. В ретикулярную формацию поступает большое количество путей из других мозговых структур. С одной стороны, это коллатерали волокон, проходящих через ствол мозга сенсорных восходящих систем, эти коллатерали заканчиваются синапсами на дендритах и соме нейронов ретикулярной формации. С другой стороны, нисходящие пути, идущие из передних отделов мозга (в том числе, пирамидный путь), тоже дают большое количество коллатералей, которые входят в ретикулярную формацию и вступают в синаптические соединения с ее нейронами. Обилие волокон поступает к нейронам ретикулярной формации из мозжечка. Таким образом, по организации своих афферентных связей эта система приспособлена к объединению влияний из различных мозговых структур. Выходящие из нее пути могут оказывать в свою очередь влияния как на вышележащие, так и на нижележащие мозговые центры.Организация межнейронных связей в ретикулярной формации достаточно дифференцирована и отдельные ее клетки соединены между собой довольно специфичными связями.
В РФ ствола мозга различают 2 отдела:
1. растральный - ретикулярная формация на уровне промежуточного мозга;
2. каудальный - ретикулярная формация продолговатого мозга, моста и среднего мозга.
Изучены 48 пар ядер РФ.
Основной функцией РФ является регуляция уровня активности коры большого мозга, мозжечка, таламуса, спинного мозга.
Характер неспецифических влияний РФ. РФ ствола мозга участвует в регуляции вегетативных функций организма. Однако еще в 1946 г. американский нейрофизиолог X.Мегоун и его сотрудники обнаружили, что ретикулярная формация имеет непосредственное отношение к регуляции соматической рефлекторной деятельности. Было доказано, что ретикулярная формация оказывает диффузное неспецифическое, нисходящее и восходящее влияние на другие мозговые структуры. Нисходящее влияние. При раздражении РФ заднего мозга (особенно гигантоклеточной ядра продолговатого мозга и ретикулярного ядра моста, где принимают начало ретикулоспинальному пути), возникает торможение всех спинальных двигательных центров (сгибательных и разгибательных). Это торможение очень глубокое и продолжительное. Такое положение в естественных условиях может наблюдаться при глубоком сне.
Наряду с диффузными тормозящими влияниями, при раздражении определенных участков РФ выявляется диффузное влияние, которое облегчает деятельность спинальной двигательной системы. РФ играет важную роль в регуляции деятельности мышечных веретен, изменяя частоту разрядов, поступающие гамма-эфферентными волокнами к мышцам. Таким образом, модулируется обратная импульсация в них.
Восходящие влияния. Наряду с функциями, которые осуществляются через нисходящие пути, у ретикулярной формации есть не менее существенные функции, которые осуществляются через ее восходящие пути. Они связаны с регуляцией активности высших отделов мозга, главным образом, коры больших полушарий. Кроме ретикулярной формации ствола мозга, в восходящую активирующую систему головного мозга входят также неспецифические ядра таламуса, задний гипоталамус, лимбических структуры. Являясь важным интегративным центром, ретикулярная формация, в свою очередь, является частью более глобальных интеграционных систем мозга, которые включают гипоталамо-лимбические и неокортикальные структуры. Именно во взаимодействии с ними и формируется целесообразное поведение, направленное на приспособление организма к меняющимся условиям внешней и внутренней среды. Таким образом, ретикулярная формация ствола головного мозга выполняет функции восходящей активирующей системы мозга, которая поддерживает на высоком уровне возбудимость нейронов коры большого мозга.
30.В состав промежуточного мозга входят: зрительные бугры (таламус), гипоталамус, эпиталамус, метаталамус. Наиболее важны таламус и гипоталамус.
Таламус - массивное парное образование, занимающее основную часть промежуточного мозга. Таламус состоит из 120 пар ядер, которые образуют 3 зоны: передняя, латеральная, медиальная. Таламус имеет 2-х сторонние связи со спинным мозгом, ретикулярной формацией ствола мозга, с гипоталамусом, с подкорковыми ядрами и корой головного мозга. В функциональном отношении ядра зрительных бугров делятся на специфические и неспецифические. Через специфические ядра проходит специфический путь передачи импульсов в кору головного мозга, т. е. к этим ядрам поступают импульсы ото всех афферентных систем (кроме обонятельной). Таким образом, зрительные бугры - коллектор афферентной информации. Специфические ядра могут изменять уровень активности коры головного мозга (модулирующее воздействие). Они играют большую роль в возникновении болевых ощущений, обеспечении эмоциональных реакций человека. Специфические ядра подразделяются на:
релейные (внешние) - получают импульсы от афферентных систем непосредственно и передают импульсы в первичные проекционные зоны (строго специфичны). За счет этих импульсов возникает ощущения. Еще импульсы идут к ассоциативным ядрам;
ассоциативные ядра (внутренние) - нет прямых контактов с афферентными системами. Получают импульсы от релейных ядер. От них импульсы идут в кору головного мозга в ассоциативные зоны (в третичные проекционные зоны), за счет этих импульсов возникают примитивные ощущения; также они обеспечивают взаимосвязь между сенсорными системами в коре головного мозга.
Неспецифические ядра зрительных бугров выполняют функции аналогичные функциям нейронов ретикулярной формации, т. е. посылают импульсы по всей коре головного мозга, оказывая стимулирующее действие.
Зрительные бугры выполняют важную функцию трансляции и обобщения сенсорных сигналов, которые поступают в мозг, являются воспринимающей областью сенсорно-двигательных импульсов. Таламус фильтрует, перерабатывает, интегрирует и направляет в мозг информацию, которая поступает от тактильных, болевых, температурных, вибрационных, мышечно-суставных, зрительных, слуховых и вкусовых путей и рецепторов, т.е в нем происходит формирование ощущений и дальнейшая их передача в кору большого мозга. Для каждого вида чувствительности существуют определенные специфические ядра (так называемая латеральная группа). В этих ядрах импульсы, идущие от органов чувств, переходят с одной нервной клетки на другую, а затем направляются в соответствующую зону мозга. Таламус является высшим центром интеграции. Таламус обеспечивает осуществление всех необходимых рефлексов, путем выполнения первичного анализа и синтеза всех возбуждений, которые поступают от рецепторов в кору головного мозга. Существует предположение, что таламус участвует в ощущении удовольствия и ощущении боли. В таламусе начинается область ретикулярной формации, которая имеет отношение к регуляции двигательной активности. Зрительные бугры также играют важную роль в контроле бодрствования и сна. Благодаря тому, что таламус осуществляет связи между корой головного мозга, с одной стороны, и базальными ганглиями и мозжечком с другой, он обеспечивает контроль сознания над автоматическими движениями. Зрительные бугры выполняют важную функцию трансляции и обобщения сенсорных сигналов, которые поступают в мозг, являются воспринимающей областью сенсорно-двигательных импульсов. Таламус фильтрует, перерабатывает, интегрирует и направляет в мозг информацию, которая поступает от тактильных, болевых, температурных, вибрационных, мышечно-суставных, зрительных, слуховых и вкусовых путей и рецепторов, т.е в нем происходит формирование ощущений и дальнейшая их передача в кору большого мозга. Для каждого вида чувствительности существуют определенные специфические ядра (так называемая латеральная группа). В этих ядрах импульсы, идущие от органов чувств, переходят с одной нервной клетки на другую, а затем направляются в соответствующую зону мозга. Таламус является высшим центром интеграции. Таламус обеспечивает осуществление всех необходимых рефлексов, путем выполнения первичного анализа и синтеза всех возбуждений, которые поступают от рецепторов в кору головного мозга. Существует предположение, что таламус участвует в ощущении удовольствия и ощущении боли. В таламусе начинается область ретикулярной формации, которая имеет отношение к регуляции двигательной активности. Зрительные бугры также играют важную роль в контроле бодрствования и сна. Благодаря тому, что таламус осуществляет связи между корой головного мозга, с одной стороны, и базальными ганглиями и мозжечком с другой, он обеспечивает контроль сознания над автоматическими движениями.
Боль — это эмоциональная реакция организма на повреждающее воздействие.Таламус участвует в передаче и переработке информации: специфические ядра таламуса обеспечивают анализ локализации болевого раздражения, его силы и длительности, неспецифические ядра таламуса участвуют в формировании мотивационно-аффективного аспекта боли.
31. Гипоталамус- высший подкорковый центр вегетативной регуляции. На висцеральные функции организма он влияет двумя путями. Во-первых через вегетативную нервную систему. Его передние ядра являются высшими парасимпатическим центрами. Поэтому при их возбуждении урежаются сердцебиения, снижается АД, понижается энергетический обмен, температура тела. суживаются зрачки и т.д.. При возбуждении задних ядер возникает обратная картина, т.к. они являются высшими симпатическими центрами.
Во-вторых. ГТ влияет на многие функции через гипофиз. Посредством нервных и сосудистых связей он образует с ним единую гипоталамо-гипофизарную систему. Нейросекреция-это способность продуцировать гормоноподобные вещества. В частности, в супраоптическом ядре вырабатываются нейрогормоны вазопрессин и окситоцин. По аксонам секретирующих нейронов они поступают в заднюю долю гипофиза, а оттуда выделяются в кровь. В медиальных ядрах синтезируются либерины и статины. По венозной гипоталамо-гипофизарной сети они транспортируются к передней доле гипофиза. В свою очередь тропные гормоны влияют на функции других желез внутренней секреции. Благодаря многочисленным связям, высокой чувствительности нейронов гипоталамуса к составу омывающей его крови, отсутствию в этом отделе гематоэнцефалического барьера, в нем находятся центры терморегуляции, регуляции водно-солевого обмена, обмена белков, жиров, углеводов и др. За счет них регулируется гомеостаз. Гипоталамус участвует в формировании некоторых мотиваций и поведенческих реакций (голода, жажды). При раздражение вентромедиального ядра чувство голода и соответствующее поведение исчезают. При его разрушении наоборот наступает неутолимый голод. Т.е. здесь находятся центры голода и насыщения. При раздражении паравентрикулярного ядра развивается чувство жажды и питьевое поведение, а при разрушении жажда исчезает. В гипоталамусе расположены центры бодрствования и сна. В опытах с самораздражением (Олдс), когда в определенные ядра ГТ вживляются электроды, установлено, что здесь находятся центры двух базисных эмоций удовольствия и неудовольствия. При раздражении некоторых ядер ГТ у человека возникает эйфория, повышается сексуальность. ГТ принадлежит важная роль в развитии стресса, т.е. реакций напряжения на угрожающую ситуацию. При воздействии физиологических или психологические стрессоров (холод, недостаток кислорода, эмоциональном напряжении) кора посылает сигналы к симпатическим центрам-ГТ, которые активируют симпатический отдел вегетативной нервной системы, выделение АКТГ, выделяются адреналин из мозгового слоя и кортикостероиды.
При патологии ГТ возникают расстройства терморегуляции (гипер- и гипотермия). аппетита (афагия-, гиперфагия), сна. Эндокринные нарушения, связанные с гипоталамусом, могут проявляться преждевременным половым созреванием, нарушениями менструального цикла, полового влечения, несахарным диабетом.
32. Подкорковые ядра-скопления серого вещества в толще нижней и боковой стенок БП. К ним относятся полосатое тело, бледный шар и ограда. Полосатое тело состоит из хвостатого ядра и скорлупы. К нему идут афферентные нервные волокна от двигательных и ассоциативных зон коры, таламуса черной субстанции среднего мозга. Связь с черной субстанцией осуществляется с помощью дофаминергических синапсов. Выделяющийся в них дофамин тормозит нейроны полосатого тела. Кроме того, сигналы к полосатому телу поступают от мозжечка, красного и вестибулярных ядер. От него аксоны нейронов идут к бледному шару. В свою очередь, от бледного шара эфферентные пути идут таламусу и двигательным ядрам среднего мозга, т.е красному ядру и черной субстанции. Полосатое тело оказывает на нейроны бледного шара преимущественно тормозящее влияние. Основная функция подкорковых ядер это регуляция движений. Кора посредством подкорковых ядер организует и регулирует дополнительные, вспомогательные движения, необходимые для правильного выполнения основного двигательного акта или облегчающие его. Это, например, определенное положение туловища и ног при выполнении работы руками. При нарушении функций подкорковых ядер вспомогательные движения становятся либо чрезмерными, либо полностью отсутствуют. В частности, при болезни Паркинсона или дрожательном параличе, полностью исчезает мимика и лицо становится маскообразным, ходьба осуществляется мелкими шажками. Больные с трудом начинают и оканчивают движение выражен тремор конечностей. Тонус мышц повышается. Возникновение болезни Паркинсона обусловлено нарушением проведения нервных импульсов от черной субстанции к полосатому телу через дофаминергические синапсы, обеспечивающие эту передачу С поражением полосатого тела и гиперактивностью бледного шара связаны заболевания с избыточными движениями. т.е. гиперкинезы. Это подергивания мыши лица, шеи, туловища, конечностей. А также двигательная гиперактивность в виде бесцельного перемещения. Например она наблюдается при хорее. Кроме этого полосатое тело принимает участие в организации условных рефлексов, процессах памяти, регуляции пищевого поведения.
33. К лимбической системе относятся обонятельные луковицы, гиппокампус, поясная извилина, зубчатая фасция, парагиппокампальная извилина, а также подкорковое миндалевидное ядро и переднее таламическое ядро. Структуры лимбической системы имеют многочисленные двусторонние связи м/у собой а также с лобными, височными долями коры и гипоталамусом благодаря этим связям она регулирует и выполняет следующие функции. 1. Регуляция вегетативных функций и поддержание гомеостаза. ЛС называют висцеральным мозгом, осуществляет тонкую регуляцию функций органов, реагирует на небольшие отклонения гомеостаза. Она влияет на эти функции через вегетативные центры гипоталамуса и гипофиз.об этом свидетельствует разнообразные изменения их функций при раздражении лимбических структур,особенно миндалин.при этом эффекты: повышение или понижение частоты сердечных сокращений, моторики и секреции желудка и кишечника,секреции различных гормонов аденогипофизом(особенно АКТГ и гонадотропинов)
2. Формирование эмоций. Раздражение миндалевидного ядра вызывает появление у пациентов беспричинных эмоций страха, гнева, ярости. При удалении миндалевидного ядра у животных. полностью исчезает агрессивное поведение. Раздражение некоторых зон поясной извилины ведет к возникновению немотивированной радости или грусти
3. Формирование мотиваций. Миндалевидное ядро регулирует пищевую мотивацию. Некоторые его области тормозят активность центра насыщения и стимулируют центр голода гипоталамуса. Другие действуют противоположным образом. За счет этих центров пищевой мотивации миндалевидного ядра формируется поведение на вкусную и невкусную пищу. В нем есть отделы, регулирующие половую мотивацию. При их раздражении возникает гиперсексуальность и выраженная половая мотивация.
4. Участие в механизмах памяти. Функция связана с основным лимбическим кругом Пейпеца
Гиппокамп классифицирует и кодирует всю информацию, которая должна быть заложена в долговременной памяти. Во-вторых обеспечивает извлечение и воспроизведение нужной информации в конкретный момент. Миндалина индуцирует сильные отрицательные эмоции,что способствует быстрому и прочному формированию временной связи.
34. В коре головного мозга выделяют: древнюю, старую и новую кору. Древняя и старая кора объединяются с некоторыми близлежащими ядрами и образуют лимбическую систему. Толщина новой коры - 3 мм, включает много извилин, площадь новой коры 2500 см2, 3 вида структур коры головного мозга: нервные клетки, отростки нервных клеток, нейроглия.
В составе коры головного мозга - различные по строению нейроны - звездчатые, большие и малые пирамидные, веретенообразные, корзинчатые и другие.
В функциональном отношении все нейроны подразделяются на:
афферентные (звездчатые клетки) - к ним идут импульсы от специфических путей и возникают специфические ощущения. Они передают импульсы к вставочным и эфферентным нейронам. Группа полисенсорных нейронов - получает импульсы от ассоциативных ядер зрительных бугров;
эфферентные нейроны (большие пирамидные клетки) - импульсы от них идут на периферию и обеспечивают определенный вид деятельности;
вставочные нейроны (малые пирамидные, веретенообразные и другие). Вставочные нейроны могут быть возбуждающими и тормозными (большие и малые корзинчатые нейроны, нейроны с кистеобразными аксонами, канделяброобразные нейроны).
Функции отростков нервных клеток:
обеспечивают связь в пределах коры головного мозга между выше- и нижележащими клетками;
обеспечивают связь в пределах одного полушария коры головного мозга;
комиссуральные - выходят из коры головного мозга, проходят через комиссуру и идут в кору головного мозга противоположного полушария;
выходят из коры головного мозга и идут в нисходящем направлении, образуя пирамидные и экстрапирамидные пути.
Нейроны и отростки в коре головного мозга расположены 6-ю слоями. Нейроны выполняющие одну и ту же рефлекторную функцию расположены строго друг над другом. Таким образом, структурной единицей коры головного мозга являются отдельные колонки. Наиболее выражена связь между 3, 4, 5 слоями коры головного мозга.
Доказательство наличия колонок: при введении микроэлектродов в кору строго перпендикулярно регистрируется импульс при воздействии одной и той же рефлекторной реакции. Если вводить электроды в горизонтальном направлении - регистрируются импульсы характерные для разных рефлекторных реакций. Диметр колонки - 500 мкм. Соседние колонки тесно связаны в функциональном отношении и часто находятся друг с другом в реципрокных взаимоотношениях (одни - возбуждают, другие - тормозят). При действии раздражителя в ответную реакцию вовлекаются много колонок, происходит совершенный анализ и синтез раздражений - принцип экранирования (при раздражении строго определенного количества рецепторов в коре головного мозга в ответную реакцию вовлекается значительная зона).
Функции коры головного мозга
Кора головного мозга - высший распорядитель и распределитель функции организма животного и человека (И. П. Павлов).
Функции коры головного мозга:
делает совершенной взаимосвязь между органами и тканями внутри организма;
обеспечивает сложные отношения организма с внешней средой;
обеспечивает процессы мышления и сознания;
является субстратом высшей нервной деятельности.
Функциональная ассиметрия полушарий является важнейшим психофизиологическим свойством головного мозга человека. При исследовании речи:словесный информационный канал контролируется левым полушарием,а несловесный(интонация)правым.абстрактное мышление и сознание связаны с левым полушарием.при вырботке условного рефлекса в начальной фазе доминирует правое полушарие,а во время упрочнения рефлекса-левое.правое полушарие осуществляет обработку информации синтетически,при этом лучше воспринимаются пространсвтенные и относительные признаки предметов.левое аналитически,лучше воспрнимая абсолютные признаки предметов и временные отношения.в эмоциональной сфере правое полушарие обуславаливает отрицательные эмоции,контролирует появление сильных эмоций,в целом более «эмоционально».левое в основном положительные эмоции контролирует проявление более слабых эмоций.
35.Вегетативная нервная система — часть нервной системы, регулирующая деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов и отчасти мускулатуры. Термин «вегетативная нервная система» ввел в 1801 г. французский врач М. Биша, отграничив тем самым вегетативную нервную систему, управляющую обменом веществ и тесно связанными с ним функциями (кровообращением, дыханием, пищеварением, выделением, размножением), от соматической нервной системы, воспринимающей раздражения из окружающей среды и координирующей реакции скелетной мускулатуры. Активность вегетативной нервной системы в значительно меньшей степени, чем соматической, воспринимается и контролируется сознанием, поэтому иногда ее называют также «автономной», или «непроизвольной», нервной системой. Вегетативная нервная система делится на два отдела — симпатический и парасимпатический. Деление это до некоторой степени условно.
Симпатические спинномозговые центры, от которых начинаются периферические симпатические волокна, расположены в боковых рогах спинного мозга (см. Центральная нервная система, спинной мозг). От скоплений симпатических клеток отходят тонкие волокна, вступающие в передние корешки спинного мозга и вместе с ними выходящие из спинного мозга. Подходя к узлу (ганглию) симпатического ствола, эти волокна вступают в него и заканчиваются на его клетках, от которых начинается новый периферический нейрон (нервная клетка с ее отростками), т. е. происходит «переключение» исходящих из центральной нервной системы предузловых (преган-глионарных) вегетативных волокон в периферических нервных узлах (ганглиях) на послеузловые (постганглионарные) нейроны, идущие к рабочему органу. Симпатические стволы, расположенные по обеим сторонам позвоночника, состоят из 2—3 шейных узлов, 12 грудных, 2—5 поясничных, 2—5 крестцовых и одного непарного — копчикового, которым замыкаются цепочки узлов симпатических стволов. Не все преганглионарные волокна заканчиваются в клетках узлов симпатического ствола, часть их не прерывается в узлах, а уходит на периферию и заканчивается в одном из предпозвоночных узлов (чревное сплетение, нижнебрыжеечное сплетение и др.). Послеузловые волокна от симпатических клеток этих узлов направляются к рабочему органу, чаще всего вместе с кровеносными сосудами в виде сплетений на них. Т. о., симпатическая иннервация внутренних органов и ряда других образований зависит от рефлекторной деятельности систем, берущих начало в боковых рогах грудного и поясничного отделов спинного мозга.
Симпатическая нервная система расширяет зрачок, вызывает учащение пульса и повышение кровяного давления, расширяет мелкие бронхи, способствует сокращению сфинктеров мочевого пузыря и прямой кишки.
Парасимпатическая иннервация осуществляется нервными центрами, находящимися в крестцовом отделе спинного мозга и в стволе головного мозга, причем первые регулируют деятельность органов, расположенных в малом тазу (мочевой пузырь, прямая кишка и половые органы), а парасимпатические центры головного отдела иннервируют остальные органы через блуждающий, языко-глоточный, глазодвигательный и другие нервы, парасимпатические ядра которых расположены в стволе головного мозга (см. Центральная нервная система, головной мозг). От головных парасимпатических центров (ядер) начинаются длинные предузловые волокна, которые подходят к рабочему органу и переключаются на послеузловые волокна уже в стенке рабочего органа во внутриорганных (внутристеночных) узлах или же (например, для слезной, слюнных желез, гладкой мускулатуры глаза) на клетках парасимпатических узлов, расположенных в непосредственной близости от иннервируемых органов (ушной, подчелюстной, ресничный и другие узлы). От крестцового парасимпатического центра предузловые волокна достигают органов малого таза в составе тазовых внутренностных нервов. Действие парасимпатической нервной системы во многом противоположно действию симпатической системы: парасимпатическая система суживает зрачок, замедляет сердечную деятельность, снижает артериальное давление. При повышении тонуса парасимпатической нервной системы отмечается наклонность к спазму мелких бронхов, учащению мочеиспускания и дефекации. Долгое время считали, что между симпатической и парасимпатической отделами вегетативной нервной системы существует антагонизм. Однако оба эти отдела совместно обеспечивают изменения жизненных функций организма и создают гомеостаз — гармоничное, физиологическое состояние органов и систем целостного организма на оптимальном уровне.
Контроль за действием симпатической и парасимпатической систем осуществляется центральными вегетативными аппаратами, расположенными в головном мозге. Сложнейшие задачи соматовегетативной интеграции (объединения в единое целое) решаются на уровне центральной нервной системы. Именно здесь, в коре больших полушарий, а также высших вегетативных центрах (гипоталамусе, лимбической системе, в среднем и продолговатом мозге) определяются изменения кровообращения, дыхания, пищеварения и т. д., без которых невозможно целостное поведение организма. В условиях целостного организма ни одно из действий вегетативной нервной системы не происходит изолированно, в отрыве от функций других отделов вегетативной нервной системы и соматической нервной системы. Благодаря соматовегетативной интеграции каждый поведенческий акт (реакция на воздействие окружающей среды) содержит соматические, симпатические и парасимпатические компоненты. Например, при оборонительной реакции, кроме активности скелетных мышц (соматическая реакция), усиливается сердечная деятельность (симпатич. реакция), расширяются сосуды функционирующих мышц (отчасти парасимпатическая реакция), а в остальных органах (внутренние органы, кожа и др.) сосуды суживаются (симпатическая реакция).
36. Большинство внутренних органов обладают двойной иннервацией: к каждому их них подходят два нерва — симпатический и парасимпатический. На многие органы симпатический и парасимпатический нервы оказывают противоположное влияние. Так, симпатический нерв ускоряет и усиливает работу сердца, а парасимпатический (блуждающий) тормозит; парасимпатический нерв вызывает сокращение кольцевой мускулатуры радужной оболочки глаза и в связи с этим сужение зрачка, а симпатический нерв вызывает расширение зрачка (сокращение радиальной мускулатуры радужной оболочки). Вместе с тем их влияние на деятельность целостного организма таково, что они могут выступать как функциональные синергисты, т.е. давать однозначный эффект. Так, в случае повышения кровяного давления возвращение его к исходному уровню может быть достигнуто как снижением активности симпатической системы, так и увеличением активности парасимпатической. Некоторые органы снабжаются волокнами только парасимпатической системы (слюнные железы, железы носоглотки, сфинктер зрачка) или симпатической (почти все кровеносные сосуды, печень, жировые клетки, половые органы, секреторные клетки поджелудочной железы). Во многих органах, иннервируемых как симпатической, так и парасимпатической системами в условии адаптированного состояния, в покое преобладают влияния парасимпатической системы. Вместе с тем их влияние на деятельность целостного организма таково, что они могут выступать как функциональные синергисты, т.е. давать однозначный эффект. Так, в случае повышения кровяного давления возвращение его к исходному уровню может быть достигнуто как снижением активности симпатической системы, так и увеличением активности парасимпатической. Некоторые органы снабжаются волокнами только парасимпатической системы (слюнные железы, железы носоглотки, сфинктер зрачка) или симпатической (почти все кровеносные сосуды, печень, жировые клетки, половые органы, секреторные клетки поджелудочной железы). Во многих органах, иннервируемых как симпатической, так и парасимпатической системами в условии адаптированного состояния, в покое преобладают влияния парасимпатической системы.
Все вегетативные рефлексы делятся на типичные и нетипичные.
I. Нетипичные рефлексы самые простые и самые содружественные. В них не участвуют центральные ЦНС. К ним относятся: 1) Аксон - рефлекс осуществляющийся в пределах ветвления постганглионарного волокна, без участия тела нейрона Дело в том. что один аксон, разветвляясь на несколько ветвей, может иннервировать много органов. Поэтому раздражение одной ветви аксона может вызвать ответную реакцию другого отдаленного от него аксона. 2) периферические вегетативные рефлексы возникают при участии клеток Догеля интрамурального ганглия. Клетки Догеля II типа воспринимают действия раздражителя и передают возбуждение на клетки I типа, которые находятся под контролем центрального отдела ВНС. Под их влиянием изменяется возбудимость этих клеток. II. Типичные рефлексы осуществляются при обязательном участии ЦНС. Они различаются в зависимости от локализации рецепторов, раздражение которых вызывает рефлекс, и эффекторов, участвующих в конечной реакции. В связи с этим различают:
1) Висцеро - висцеральные рефлексы.
Они возникают с рецепторов внутренних органов и влияют на деятельность этих органов. Внсцеро-внсцеральные рефлексы бывают двух видов: собственные и сопряженные.
Если рефлексы возникают и осуществляются в одной системе, то это собственные висцеро-висцеральные рефлексы. Так, при изменении давления крови, изменяется деятельность сердца. Это собственный рефлекс, так как рецепторы и эффектор относятся к системе кровообращения.
Сопряженные висцеро-висцеральные рефлексы возникают в одной системе, а на другую влияют Так. при раздражении органов брюшной полости может произойти остановка сердца, при холецистите наблюдаются роли в сердце.
2)Висцеро - дермальиые рефлексы.
Они возникают с интерорецепторов, а проявляются в изменении потоотделения, кожной чувствительности и т. д. на поверхности тела. Так, при стенокардии часто возникают боли на поверхности тела в области руки, лопатки, уха и т. д.
3) Висцеро-моторные рефлексы.
Они возникают с интерорецепторов и изменяют тонус мышц. Так при аппендиците повышается тонус напряжение мышц в правой подвздошной области.
4) Дермо - висцеральные рефлексы.
Они проявляются в том, что при раздражении некоторых участков кожи наступают сосудистые реакции и другие изменения деятельности внутренних органов. На этом основано применение ряда лечебных процедур: банки, парафин, горчичники, рефлексотерапия.
Вегетативные рефлексы возникают и при возбуждении проприорецепторов. Вегетативные компоненты являются обязательным сопровождением любого поведенческого акта. Самые различные акты поведения, проявляющиеся в мышечной деятельности, в активных движениях, всегда сопровождаются изменение внутренних органов, органов кровообращения, пищеварения, выделения, внутренней секреции. Благодаря изменениям ФУНКЦИИ внутренних органов обеспечивается максимально-оптимальная, эффективная и длительная работа мышц, которая возможна лишь при увеличении кровоснабжения мышц и усилении в них обмена веществ.
37.Практически все сенсорные приборы на периферии имеют вспомогательные структуры для восприятия раздражающего стимула. Эти вспомогательные структуры обеспечивают лучшее восприятие стимулов и поэтому играют важную роль в деятельности сенсорных систем.
Вспомогательные структуры могут быть очень сложными. Например, оптическая система глаза, или более простыми - тактильные рецепторы в коже. Через вспомогательные структуры внешний стимул доходит до рецептирующего субстрата, определяющего модальность рецептора, и взаимодействует с ним.
Рецептор - это специализированная структура (клетка или окончание афферентного волокна), которая в процессе эволюции приспособилась к восприятию соответствующего раздражителя внешнего или внутреннего мира. Стимул, который является эффективным в данном случае, называют адекватными раздражителем. В физиологических исследованиях было обнаружено, что каждый рецептор реагирует чаще всего на стимулы какого-то одного типа (мономодальный рецептор). Рецепторы на основе их активации адекватными стимулами делят на четыре основные группы: механо-, термо-, хемо- и фоторецепторы. Причем, внутри каждой из этих групп можно выделить свои подгруппы.
Таким образом, все рецепторы любой сенсорной системы являются обязательной частью периферического отдела анализатора С периферии информация передается в ЦНС в закодированной форме, в виде потенциалов действия (нервных импульсов) по афферентным нервным волокнам.
К проводниковому отделу каждого анализатора относятся афферентные нервные волокна и нервные клетки спинного, продолговатого и промежуточного отделов мозга, аксоны которых проводят импульсы к коре боль- ших полушарий.
Афферентные нейроны - это первые нейроны, которые участвуют в обработке сенсорной информации, поступающей от рецепторов кожи, мышц, суставов и внутренних органов. Как правило, тела афферентных нейронов лежат в различных ганглиях (спинномозговых, головы и шеи. вестибулярном, спиральном, коленчатом и т.д.), за исключением зрительного анализатора, афферентные нейроны которого (ганглиозные клетки) находятся непосредственно на сетчатке.
От всех частей тела, за исключением головы, периферические нервы (афферентные волокна) идут к спинномозговым ганглиям и аксоны этих клеток в составе задних корешков поступают в спинной мозг.
Таким образом, основная функция афферентных нервных волокон - это передача информации с рецепторного аппарата.
Второй нейрон, участвующий в обработке информации, может быть расположен в спинном или продолговатом мозге. Отсюда импульсы через средний мозг идут к ядрам таламуса. в которых располагается у большинства анализаторов следующий (предпоследний) нейрон, участвующий в обработке сенсорной информации. Исключением из этого правила является обонятельный анализатор - после обонятельной луковицы информация направляется в обонятельную кору, не заходя в таламус.
От общего сенсорного коллектора (таламуса) информация поступает в центральный отдел анализатора. Для каждого анализатора имеются свои соответствующие проекционные и ассоциативные зоны коры, куда приходят импульсы от рецепторного аппарата. В проекционных зонах происходит декодирование информации, возникает представление о модальности сигнала, о его силе и качестве, а в ассоциативных участках коры происходит определение «что это такое?» - акцепция сигнала. Это происходит с участием процессов памяти. Центральный отдел так же контролирует процесс обработки информации в нижележащих структурах мозга. Например, усиление чувствительности фоторецепторов в условиях темновой адаптации.
38. Зрительный анализатор является сложной нервно-рецепторной системой, предназначенной для восприятия и анализа световых раздражений. В нем, как и в любом анализаторе, имеются три основных отдела - периферический, проводниковый и центральный.
Периферический отдел зрительного анализатора особенно сложен. Он представлен глазным яблоком, которое содержит не только рецепторы, но и специфический светопреломляющий аппарат (роговицу, жидкость передней камеры глаза, хрусталик и стекловидное тело). Преломляющую силу оптической системы выражают в диоптриях. Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила здорового глаза составляет 59 диоптрий (при рассматривании далеких предметов) и 70,5 диоптрий (при рассматривании близких предметов).
Неодинаковая кривизна преломляющих поверхностей оптической системы глаза в горизонтальном и вертикальном меридианах (чаще всего роговой оболочки) приводит к развитию астигматизма. Изменение длины глазного яблока приводит к развитию миопии (близорукости) или гиперметропия (дальнозоркости). Снижение эластических свойств хрусталика – к развитию пресбиопии (старческой дальнозоркости).
Имеющаяся радужная оболочка регулирует интенсивность светового потока. Заложенные в ней мышцы имеют различную иннервацию. Радиальные мышцы иннервируются симпатическими нервами, поэтому при активации симпатического отдела автономной нервной системы зрачок расширяется. Кольцевые мышцы иннервируются парасимпатическим отделом и при его возбуждении диаметр зрачка уменьшается.
Анализатор зрения состоит из периферической части – орган зрения, проводящий путь – зрительный нерв, центральная часть-затылочная доля кбпгм Орган зрения состоит из глазного яблока и вспомогательных образований (мышцы, веки, слезный аппарат) Глазное яблоко состоит из ядра, образованного 3 оболочками: фиброзной, сосудистой и внутренней сетчатки. Фиброзная оболочка делится на склеру и роговицу (радужная оболочка) Сосудистая состоит из собственно сосудов и реснитчатого тела (продолжение сосудистой оболочки и прикрепляется к хрусталику) Сетчатка состоит из 2 листков – внутреннего (светочувствительного) и наружного (пигментного). Кроме этого внутренний листок сетчатки делится на 3 слоя : 1. наружный (фоторецепторный слой, периферическими отростками которого являются палочки и колбочки) 2. ассоциативный 3. внутренний (ганглионарный) Палочки – образования, которые воспринимают информация о освещенности и форме предметов и обеспечивают ночное видение. В сетчатки человека содержится 1 вид палочек. Колбочки – образования, которые воспринимают цветную информацию и обеспечивают дневное видение. Содержится 3 типа колбочек Хрусталик – образование, представляющую собой абсолютно прозрачную, двояковыпуклую линзу (диа. 9 мм) , соединена Цинной связкой с ресничной мышцей (ресничным телом). Аккомодация – изменение выпуклости хрусталика. Причина – расслабление или сокращение ресничной мышцы, при этом расслабление приводит к уплощению хрусталика и обеспечивает видеть предметы, расположенные вдали. Сокращение приводит к увеличению выпуклости хрусталика и обеспечивает возможность видеть предметы вблизи. Стекловидное тело – (межклеточное или желеобразное вещество) – образование, заполняющие пространство между сетчаткой и хрусталиком, выполняющие роль линзы.
39. Аккомодация глаза — способность ясно видеть предметы, находящиеся на различных расстояниях от глаза. Физиологический механизм аккомодации глаза состоит в том, что при сокращении волокон цилиарной мышцы глаза происходит расслабление цинновой связки, при помощи которой хрусталик прикреплен к цилиарному телу При этом уменьшается натяжение сумки хрусталика, и он благодаря эластическим свойствам становится более выпуклым. Расслабление цилиарной мышцы ведет к уплощению хрусталика. Иннервация цилиарной мышцы осуществляется глазодвигательным и симпатическим нервами. Аккомодация глаза возможна в пределах, ограниченных ближайшей и дальнейшей точками ясного зрения. Первая определяется наименьшим расстоянием, на котором возможно читать мелкий шрифт; вторая — наибольшим расстоянием, на котором ясно различим предмет при отсутствии аккомодации глаза. Положение дальнейшей точки ясного зрения зависит от рефракции глаза . Увеличение преломляющей силы оптической системы глаза, достигаемое при максимальном напряжении аккомодации глаза, называют объемом, или силой, аккомодацией глаза.Объем аккомодации глаза изменяется с возрастом вследствие уменьшения эластичности хрусталика К патологическим изменениям относят спазм, паралич и парез аккомодации глаза. Спазм возникает обычно у молодых людей при длительном напряжении аккомодации глаза, травме, действии на глаз очень яркого света. Спазм аккомодации глаза проявляется близорукостью. Параличи и парезы аккомодации глаза могут быть центрального происхождения и обусловливаться инфекциями и интоксикациями. Периферические параличи аккомодации глаза наблюдают при ушибах глаза, приеме внутрь препаратов атропина, при закапывании в конъюнктивальный мешок средств, расширяющих зрачок. Паралич аккомодации глаза характеризуется невозможностью различать мелкий шрифт на близком расстоянии. Для лечения спазма и паралича аккомодации глаза больные подлежат направлению к врачу -окулисту.
Аккомодация глаза— свойство глаза изменять преломляющую силу для приспособления к восприятию предметов, находящихся от него на различных расстояниях. Механизм аккомодации глаза заключается в следующем: при сокращении волокон заложенной в цилиарном теле аккомодационной мышцы происходит расслабление цинновой связки, посредством которой хрусталик подвешен к цилиарному телу; в результате этого хрусталик, обладающий эластическими свойствами, приобретает более выпуклую форму, и преломляющая способность глаза усиливается . При расслаблении аккомодационной мышцы волокна цинновой связки натягиваются, хрусталик уплощается, и преломляющая сила оптической системы глаза соответственно уменьшается. Аккомодация глаза может осуществляться в определенных пределах, зависящих главным образом от эластических свойств хрусталика.
Близорукость, или Миопия — это дефект (аномалия рефракции) зрения, при котором изображение падает не на сетчатку глаза, а перед ней из-за того, что глаз слишком сильно фокусирует (относительно данного передне-заднего размера глазного яблока). Человек при этом хорошо видит вблизи, но плохо видит вдаль и должен пользоваться очками/линзами с минусовыми диоптриями.причины:наследственность, перенапряжение глаз,неправильная коррекция
Дальнозоркость,(гиперметропия) — особенность рефракции глаза, состоящая в том, что изображения далеких предметов в покое аккомодации фокусируются за сетчаткой. В молодом возрасте при не слишком высокой дальнозоркости с помощью напряжения аккомодации можно сфокусировать изображение на сетчатке.
Для получения отчетливого изображения на сетчатке приходится усилить рефракцию. Это аномалия зрения, которую имеют около четверти населения Земли. Существует ошибочное мнение, что дальнозоркие хорошо видят вдаль, однако это не всегда так. Часто дальнозоркие видят плохо и вблизи, и вдали. Однако люди, страдающие лишь возрастной дальнозоркостью, хорошо видят вдаль, т. к. у них нет аномалий рефракции, однако хрусталик находится в расслабленном состоянии. Дальнозоркие люди часто испытывают головные боли при выполнении работы вблизи. Пресбиопия - состояние глаз, которое возникает у всех без исключения (!) людей с возрастом (обычно после 40 лет). Человеку становится сложно различать мелкие предметы вблизи, читать газетный шрифт и т.п. Это происходит из-за того, что хрусталик глаза со временем становится все более плотным и все менее эластичным. Ослабевают из-за возрастных изменений мышцы, удерживающие хрусталик. Затылочные доли головного мозга, ответственные за зрение, посылают мышцам глаза сигнал, но они уже не способны в достаточной степени изменять форму хрусталика, чтобы сфокусировать изображение близко расположенных предметов на сетчатку. Дальнозоркие люди начинают испытывать проблемы с пресбиопией раньше, чем все остальные.
40. Анализатор слуха.Основной функцией органов слуха является восприятие колебаний воздушной среды. Органы слуха тесно связаны с органами равновесия. Рецепторные аппараты слуховой и вестибулярной системы расположены во внутреннем ухе.
Оба рецепторных аппарата иннервируются волокнами третьей пары черепных нервов, оба реагируют на физические показатели: вестибулярный аппарат воспринимает угловые ускорения, слуховой – воздушные колебания.
Орган слуха. Орган слуха человека состоит из наружного уха, среднего уха и внутреннего уха. Наружное ухо служит для улавливания звуков, его образуют ушная раковина и наружный слуховой проход. Ушная раковина образована эластическим хрящом, снаружи покрытым кожей. Внизу ушная раковина дополнена кожной складкой – мочкой, которая заполнена жировой тканью. Определение направления звука у человека связано с бинауральным слухом, т. е. со слышанием двумя ушами. Любой боковой звук поступает в одно ухо раньше, чем в другое. Разница во времени (несколько долей миллисекунды) прихода звуковых волн, воспринимаемых левым и правым ухом, дает возможность определить направление звука. При поражении одного уха человек определяет направление звука вращением головы.
Наружный слуховой проход у взрослого человека имеет длину 2,5 см, емкость – 1 куб. см. Кожа, выстилающая слуховой проход, имеет тонкие волоски и видоизмененные потовые железы, вырабатывающие ушную серу. Они выполняют защитную роль. Ушная сера состоит из жировых клеток, содержащих пигмент.
Наружное и среднее ухо разделяются барабанной перепонкой, представляющей собой тонкую соединительно-тканную пластинку. Толщина барабанной перепонки – около 0,1 мм, снаружи она покрыта эпителием, а изнутри – слизистой оболочкой. Барабанная перепонка располагается наклонно и начинает колебаться при попадании на нее звуковых волн. Поскольку барабанная перепонка не имеет собственного периода колебаний, то она колеблется при любом звуке соответственно его длине волны.
Среднее ухо представляет собой барабанную полость, которая имеет форму маленького плоского барабана с туго натянутой колеблющейся перепонкой и слуховой трубой. В полости среднего уха находятся сочленяющиеся между собой слуховые косточки – молоточек, наковальня и стремечко. Рукоятка молоточка вплетена в барабанную перепонку; другим концом молоточек соединен с наковальней, а последняя с помощью сустава подвижно сочленена со стремечком. К стремечку прикреплена стременная мышца, которая удерживает его у перепонки овального окна, отделяющего внутреннее ухо от среднего. Функцией слуховых косточек является обеспечение увеличения давления звуковой волны при передаче с барабанной перепонки на перепонку овального окна. Это увеличение (примерно в 30–40 раз) помогает слабым звуковым волнам, падающим на барабанную перепонку, преодолеть сопротивление мембраны овального окна и передать колебания во внутреннее ухо, трансформируясь там в колебания эндолимфы.
Барабанная полость соединена с носоглоткой при помощи слуховой (евстахиевой) трубы длиной 3,5 см, очень узкой (2 мм), поддерживающей одинаковое давление снаружи и изнутри на барабанную перепонку, обеспечивая тем самым наиболее благоприятные условия для ее колебания. Отверстие трубы в глотке чаще всего находится в спавшемся состоянии, и воздух проходит в барабанную полость во время акта глотания и зевания.
Внутреннее ухо находится в каменистой части височной кости и представляет собой костный лабиринт, внутри которого есть перепончатый лабиринт из соединительной ткани, который как бы вставлен в костный лабиринт и повторяет его форму. Между костным и перепончатым лабиринтами имеется жидкость – перилимфа, а внутри перепончатого лабиринта – эндолимфа. Кроме овального окошка, в стенке, отделяющей среднее ухо от внутреннего, есть круглое окно, которое делает возможным колебание жидкости.
Костный лабиринт состоит из трех частей: в центре находится преддверие, спереди от него – улитка, а сзади – полукружные каналы. Костная улитка – спирально извивающийся канал, образующий два с половиной оборота вокруг стержня конической формы. Диаметр костного канала у основания улитки – 0,04 мм, на вершине – 0,5 мм. От стержня отходит костная спиральная пластинка, которая делит полость канала на две части – лестницы.
Внутри среднего канала улитки находится спиральный (кортиев) орган. Он имеет базилярную (основную) пластинку, состоящую примерно из 24 тыс. тонких фиброзных волоконец различной длины. Эти волоконца очень упругие и слабо связаны друг с другом. На основной пластинке вдоль нее в пять рядов располагаются опорные и волосковые чувствительные клетки – это и есть слуховые рецепторы.
Внутренние волосковые клетки расположены в один ряд, по всей длине перепончатого канала их насчитывается 3,5 тыс. Наружные волосковые клетки располагаются в три-четыре ряда, их насчитывается 12–20 тыс. Каждая рецепторная клетка имеет удлиненную форму, на ней имеется 60–70 мельчайших волосков (длиной 4–5 мкм). Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной пластинкой, которая нависает над ними. Волосковые клетки охватываются нервными волокнами улитковой ветви слухового нерва. В продолговатом мозге находится второй нейрон слухового пути; потом путь идет, перекрещиваясь, к задним буграм четверохолмия, а от них – в височную область коры, где располагается центральная часть слухового анализатора.
В коре больших полушарий находится несколько слуховых центров. Некоторые из них (нижние височные извилины) предназначены для восприятия более простых звуков – тонов и шумов. Другие связаны со сложнейшими звуковыми ощущениями, которые возникают в то время, когда человек говорит сам, слушает речь или музыку.
41. Анализатор вкуса,обоняния. Способность различать запахи обеспечивает обонятельный анализатор. Он относится к приборам дистантного действия и состоит из воспринимающего (рецепторного) аппарата, проводящих путей и отдела головного мозга, где осуществляется высший анализ и синтез информации о запахах. Воспринимающий аппарат анализатора располагается в начальном отделе воздухоносных путей — в носовой полости.
Носовая полость разделена носовой перегородкой на две половины, каждая из которых через задние носовые отверстия (хоаны) сообщается с носоглоткой. На боковых стенках полости носа имеются выступы — носовые раковины. Это тонкие покрытые слизистой оболочкой костные пластинки, которые, располагаясь одна над другой, образуют нижний, средний и верхний носовые ходы. Обонятельная область находится в верхнем отделе полости носа и занимает площадь примерно в один квадратный сантиметр. Слизистая оболочка этой области отличается от слизистой остальной поверхности полости носа и цветом (он желтовато-коричневый, а не розовый, как повсюду) и особенно строением.
В этом месте в толще слизистой оболочки залегают обонятельные рецепторные клетки, чередующиеся с опорными; они достаточно плотно прилежат друг к другу, образуя обонятельный эпителий. Здесь же находятся и мелкие железки, выделяющие так называемую обонятельную слизь. Она увлажняет и защищает обонятельный эпителий и, кроме того, служит своеобразным фильтром для пахучих молекул. Процесс восприятия запаха начинается с рецепторной обонятельной клетки, по форме напоминающей веретено с двумя отростками: один — короткий, периферический — направляется к поверхности слизистой оболочки, другой — длинный, центральный — в головной мозг.
Периферические отростки имеют на конце утолщение в виде булавы с 10—12 тонкими волосками — ресничками. Реснички эти чрезвычайно подвижны: они сгибаются, выпрямляются, поворачиваются в разные стороны, как бы отыскивая и улавливая молекулы пахучих веществ. На обонятельных ресничках обнаружены рецептивные участки, отличающиеся особым строением и свойствами, благодаря чему они контактируют только с определенными пахучими молекулами. В результате такого контакта в рецепторной клетке рождается нервный импульс, который устремляется по центральному отростку в головной мозг.
Центральные отростки обонятельных клеток собираются в 15—20 стволиков и через мелкие отверстия решетчатой пластинки в верхних отделах полости носа уходят в полость черепа, достигая следующего отдела обонятельного анализатора — обонятельной луковицы (9). Их две, и лежат они на нижней поверхности правой и левой лобных долей коры головного мозга.
Обонятельная луковица представляет собой сложно организованный нервный центр, где осуществляется предварительная обработка всей поступающей информации о запахах. Из луковиц по двум обонятельным трактам, которые лежат параллельно средней линии черепа, сигналы поступают в кору нижней поверхности височной доли головного мозга (10), где располагается высший отдел обонятельного анализатора и после окончательной обработки и синтеза информации формируется ощущение того или иного запаха.
Вкусовой анализатор служит для определения характера, вкусовых качеств корма, его пригодности к поеданию. Животным, живущим в воде вкусовой и обонятельный анализаторы помогают ориентироваться в окружающей среде, определять наличие пищи, самки. С переходом к жизни в воздушной среде значение вкусового анализатора уменьшается. У травоядных животных вкусовой анализатор развит хорошо, что бывает видно на пастбище и в кормушке, когда животные не всю подряд поедают траву и сено.
Периферический отдел вкусового анализатора представлен вкусовыми луковицами, расположенными на языке, мягком небе, задней стенке глотки, миндалинах и надгортаннике. Вкусовые луковицы расположены на поверхности грибовидных, листовидных и желобовидных сосочков.
Луковица состоит из опорных и 2-6 рецепторных клеток с микроресничками на их поверхности. Суженая часть луковицы имеет небольшое отверстие - вкусовую пору, через которую проникает растворенное вещество, действующее на вкусовые рецепторы. Больше всего рецепторов на кончике, краях и задней части языка.
В слизистой оболочке языка и других участках ротовой полости имеются рецепторы, воспринимающие температуру, прикосновение, давление, боль. И то, что мы называем вкусом является результатом раздражения не только вкусовых, но и ряда других перечисленных в.ч. обонятельных, зрительных и др. рецепторов. Поэтому неодинаково воспринимается вкус одного и того же теплого или холодного, жидкого или более густого продукта. Временная потеря обоняния наблюдается и при насморке, что приводит к нарушению вкусовых ощущений.
Различают четыре основных вкусовых ощущения - горькое, сладкие, кислое и соленое.
Ощущение сладкого больше воспринимается передней частью языка, горького - его основанием, кислого - средней частью его боковой поверхности, соленого - кончиком и боковым краем.
Жвачные животные, лошади, свиньи хорошо различают все четыре вкуса. Однако свиньи отдают предпочтение сладкому, крупный рогатый скот и лошади - соленому.
Чувствительность вкусового анализатора в значительной степени определяется степенью потребности животного в корме, функциональным состоянием органов пищеварения и других систем организма, полноценностью и набором кормов в рационе.
Афферентные вкусовые волокна идут в составе лицевого, языкоглоточного и блуждающего нервов в продолговатый мозг, далее в таламус и в корковую зону вкусового анализатора.
42Вестибулярный анализатор обеспечивает ориентацию в пространстве: восприятие действия на организм силы земного притяжения, положения тела в пространстве, характера перемещения тела (ускорение, замедление, вращение). При любом изменении положения тела или головы в пространстве раздражаются рецепторы органа равновесия, возникший нервный импульс проводится по вестибулярному нерву в составе преддверно-улиткового нерва в головной мозг: средний мозг, мозжечок, таламус и, наконец, в кору теменной доли.
Строение и функции органа равновесия
(ВЕСТИБУЛЯРНЫЙ АППАРАТ)
Орган равновесия является частью внутреннего уха и вместе с улиткой заключен в костный лабиринт височной кости. Он представлен:
преддверием внутреннего уха с двумя расширениями - овальным и округлым мешочками
тремя полукружными каналами. Округлый и овальный мешочки и полукружные каналы заполнены жидкостью - эндолимфой.
Внутренняя поверхность мешочков образована слоем эпителиальных клеток, среди которых имеютсячувствительные волосковые клетки с тонкими чувствительными выростами. Чувствительные отростки рецепторных клеток погружены в тонкий слой студенистой массы, в которой лежит большое количество очень мелких кристалликов углекислого кальция - статолитов. Любые изменения тела или головы в пространстве, вибрационные воздействия, ускорение или замедление прямолинейного движения вызывают перемещение статолитов. При этом статолиты раздражают определенные группы рецепторных клеток, в результате человек получает сигнал об изменении положения тела.
Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях. Участки полукружных каналов, обращенные к преддверию, имеют расширения - ампулы. На внутренней поверхности ампул также имеются рецепторные клетки с чувствительными волосками, и они также погружены в тонкий слой студенистой жидкости, лежащий по внутренней поверхности ампул. Рецепторные клетки ампул тонко реагируют на малейшие перемещения эндолимфы и студенистой жидкости полукружных каналов. Перемещения жидкости возникают в результате перемещения тела или головы: ускорения, замедления движения и вращательные движения. Поскольку полукружные каналы ориентированы в трех взаимно перпендикулярных плоскостях, то любой по ворот головы или тела воспринимается вестибулярными рецепторами.Таким образом, работа вестибулярного анализатора позволяет постоянно оценивать положение и движение тела в пространстве и в соответствии с этим рефлекторно изменять тонус скелетных мышц, в необходимом направлении менять положение головы и тела.
При повреждении вестибулярного аппарата возникают головокружения, нарушается равновесие, проявляются симптомы морской болезни.
У человека чувство равновесия и оценка положения тела в пространстве связано не только с органом равновесия, но и с наличием большого количества рецепторов (барорецепторов) в мышцах и коже, которые воспринимают механическое давление на них.
43.Кровь как функциональная система. Как известно, под системой принято понимать упорядоченное целостное множествовзаимосвязанных элементов, обладающее собственной организацией и структурой. Главным свойством системы как единой совокупности взаимодействующих элементов является целостность, выражающаяся в несводимости свойств системы к сумме свойств составляющих ее частей.
В 1939 г. Г.Ф. Ланг предложил объединить кровь, регулирующий нейрогуморалъный аппарат, и органы, в которых происходит образование клеток крови и их разрушение, — костный мозг, вилочковую железу, лимфатические узлы, селезенку и печень — ввиду тесной их связи под общим понятием система крови. Компоненты этой системы осуществляют непосредственный контакт с кровяным руслом. Такое взаимоотношение обеспечивает не только транспорт клеток, но и поступление различных гуморальных факторов из крови в кроветворные органы.
Главным местом образования клеток крови у высших позвоночных является костный мозг. Здесь находится основная масса кроветворных элементов. В нем же осуществляются и разрушение эритроцитов, реутилизация железа, синтез гемоглобина, накопление резервных липидов. С костным мозгом связано происхождение популяции В-лимфоцитов, осуществляющих гуморальные реакции иммунитета, т.е. выработку антител.
Центральным органом иммуногенеза является вилочковая железа. В ней происходит образование Т-лимфоцитов, которые участвует в клеточных реакциях иммунитета, направленных на отторжение ткани. Кроме вилочковой железы ответственными за выработку иммунитета являются селезенка и лимфатические узлы. Селезенка участвует в лимфоцитопоэзе, синтезе иммуноглобулинов, разрушении эритроцитов, лейкоцитов, тромбоцитов, в депонировании крови. Лимфатические узлы продуцируют и депонируют лимфоциты.
В регуляции деятельности системы крови важную роль играют гуморальные факторы — эритропоэтины, лейкопоэтины, тромбопластины. Кроме них действуют и другие гуморальные агенты, например андрогены. Медиаторы (ацетилхолин, адреналин) влияют на систему крови не только вызывая перераспределение форменных элементов, но и путем прямого влияния на холино- и адренорецепторы клеток. Определенное влияние оказывает нервная система. Работами В.Н. Черниговского и А.Я. Ярошевского показана важная роль нервных факторов в регуляции деятельности системы крови.
Общее количество крови у взрослого человека 4—6 л, что составляет 6— 8 % массы тела (у мужчин в среднем около 5,4 л, у женщин — 4,5 л). Около 84 % крови находится в сосудах большого круга кровообращения, 9 % — малого и 7 % — в сердце. Примерно 64 % общего количества крови находится в венах, 6 % — в капиллярных и 18 % — в артериях.
Плазма крови - жидкая часть крови. В плазме крови находятся её форменные элементы (эритроциты, лейкоциты, тромбоциты). Представляет собой коллоидный раствор белков и др. органических и неорганических соединений, содержит более 20 витаминов и 20 микроэлементов (железо, фосфор, кальций, цинк, кобальт и др.).
Состав плазмы и интерстициальной жидкости существенно различается лишь по концентрации белков, так как их крупные молекулы не могут свободно проходить через стенки капилляров.
44.Кроветворение происходит непрерывно в красном костном мозге. В среднем у человека в течение жизни образуется около 450 кг эритроцитов, 5400 кг гранулоцитов, 275 кг лимфоцитов и 40 кг тромбоцитов.
Кроворазрушение также протекает непрерывно в самом сосудистом русле, в селезенке и печени в количествах, эквивалентных вновь образующимся форменным элементам.
Синтез белков плазмы происходит непрерывно в печени.
Дыхательная функция крови заключается в транспорте газов — кислорода от легких к тканям в составе артериальной крови и двуокиси углерода в обратном направлении в составе венозной крови. Основные «участники» этих процессов — сложный хромопротеид эритроцитов — гемоглобин и «летучая» угольная кислота плазмы, образующаяся в процессе метаболизма практически во всех тканях при гидратации СО2 и также легко отдающая его в лёгких.
Трофическая функция крови по отношению к клеткам заключается в переносе к ним от кишечника питательных веществ — аминокислот, липидов,
моно- и дисахаридов, витаминов, микроэлементов и др.
Экскреторная функция крови способствует выведению через почки, легкие, потовые железы и пищеварительный тракт токсичных продуктов метаболизма (мочевина, аммиак, билирубин, уробилин, двуокись углерода и др.), а также избытка воды и солей.
Защитная функция — одна из важнейших функций крови — реализуется в двух формах — иммунных реакциях (гуморальный и клеточный иммунитет) и свертывании (тромбоцитарный и коагуляционный гемостаз). Частным случаем защитной функции являются противосвертывающие механизмы системы крови.
45Эритроциты (Э)- это высоко специализированные безъядерные клетки крови. Ядро у них утрачивается в процессе созревания. Эритроциты имеют форму двояковогнутого диска. В среднем их диаметр около 7,5 мкм, а толщина на периферии 2,5 мм. Благодаря такой форме увеличивается поверхность эритроцитов для диффузии газов. Кроме того, возрастает их пластичность, за счет высокой пластичности, они деформируются и легко проходят по капиллярам. У старых и патологических эритроцитов пластичность низкая. Поэтому они задерживаются в капиллярах ретикулярной ткани селезенки и разрушаются там. Мембрана эритроцитов и отсутствие ядра обеспечивают их главную функцию - перенос кислорода и участие в переносе углекислого газа. Мембрана эритроцитов непроницаема для катионов, кроме калия, а ее проницаемость для анионов хлора, гидрокарбонат анионов и гидроксил анионов в миллион раз больше. Кроме того она хорошо пропускает молекулы кислорода и углекислого газа. В мембране содержится до 52% белка. В частности, гликопротеины определяют групповую принадлежность крови и обеспечивают ее отрицательный заряд. В нее встроена Na/К-АТФаза, удаляющая из цитоплазмы натрий и закачивающая ионы калия. Основную массу эритроцитов составляет хемопротеин гемоглобин. Кроме того, в цитоплазме содержатся ферменты карбоангидраза, фосфатазы, холинестераза и другие ферменты. Функции эритроцитов:
1. Перенос кислорода от легких к тканям.
2. Участие в транспорте СОз от тканей к легким.
3. Транспорт воды от тканей к легким, где она выделяется, в виде пара.
4. Участвуют в свертывании крови, выделяя зритроцитарные факторы свертывания.
5. Переносят аминокислоты на своей поверхности.
6.'Участвуют в регуляция вязкости крови, вследствие пластичности. В результате их способности к деформации, вязкость крови в мелких сосудах меньше, чем крупных.
В одном микро литре крови мужчин содержится 4,5-5,0млн. эритроцитов (4,5-5,0 * 1012 л). Женщин - 3,7-4,7 млн. (3.7-4.7 • 1012 л). Подсчет количества эритроцитов производится в камере Горяева. Для этого кровь в специальном капилляре меланжере (смесителе) для эритроцитов смешивают с 3% раствором хлорида натрия в соотношении 1:100 или 1:200. Затем капелька этой смеси помещается в счетную камеру. Она создается средним выступом камеры и покровным стеклом. Высота камеры 0,1 мм. На среднем выступе нанесена сетка, образующая большие квадраты. Часть этих квадратов разделена на 16 маленьких (табл.). Каждая сторона малого квадрата имеет величину 0.05мм. Следовательно, объем смеси над малым квадратом будет составлять 1/10 мм * 1/20 мм * 1/20 мм = 1/4000 мм3.
После заполнения камеры, под микроскопом считают количество эритроцитов в 5-ти тех больших квадратах, которые разделены на маленькие. Т.е. в 80 маленьких. Затем рассчитывают количество эритроцитов в одном микролитре крови по формуле:X=4000*200*(100)\80
Где а - общее количество эритроцитов, полученное при подсчете б - число малых квадратов в которых производился подсчет (80) в - разведение крови (1:100, 1:200). 4000 - величина обратная объему жидкости на малым квадратом.
Для быстрого подсчета, при большом количестве анализов, используют фотоэлектрические эритрогемометры. Принцип их действия основан на определении прозрачности взвеси эритроцитов с помощью пучка света проходящего от источника к светочувствительному датчику Фотоэлектрокалориметру. Увеличение содержания эритроцитов в крови называется эритроцитозом или эритремиеи, уменьшаться эритромении или анемией. Эти изменения могут быть относительными и абсолютными. Например, относительное уменьшение их количества возникает при задержке воды в организме а, увеличение при обезвоживании. Абсолютное уменьшение содержания эритроцитов, т.е. анемия наблюдается при кровопотере, нарушениях кроветворение, разрушении эритроцитов гемолитическими ядами или при переливании несовместимой крови.
46.Гемоглобин (НЬ) это хемопротеин, содержащийся в эритроцитах. Молекулу гемоглобина образуют четыре субъединицы, каждая из которых включает гем, соединенный с атомом железа, и белковую часть глобин. Гем синтезируется в митохондриях эритробластов, а глобин в их рибосомах. У взрослого человека гемоглобин содержит две а- и две р- полипептидных цепи. Он называется А-гемоглобнном (аdult) (-взрослый). В зрелом возрасте он составляет основную часть гемоглобина. В первые три месяца внутриутробного развития в эритроцитах находится гемоглобин типа G1 и G2 (Gоvег). В последующие периоды внутриутробного развития и в первые месяцы после рождения основную часть составляет фетальный гемоглобин (F-гемоглобин). В его структуре две а- и две у-полипептидные цепи. При рождении до 50-30% гемоглобина составляет F-гемоглобин, а 20-40 % А-гемоглобин. Ранние гемоглобины имеют большую кислородную емкость.
Гем содержит атом 2-х валентного железа, который легко соединяется с кислородом и легко отдает его. При этом валентность железа не изменяется. Один грамм гемоглобина способен связывать 1,34 мл кислорода. Соединение гемоглобина с кислородом, образующееся в капиллярах легких называется оксигемоглобином (НЬС>2). Он имеет ярко алый цвет. Гемоглобин, отдавший кислород в капиллярах тканей, называется дезоксигемоглобином или
восстановленным (НЬ). У него темно-вишневая окраска. От 10 до 30% углекислого газа, поступающего из тканей в кровь, соединяются с амидной группировкой гемоглобина. Образуется легко диссоциирующее соединение карбгемоглобин (НЪС0I). В этом виде часть углекислого газа транспортируется к легким.
В некоторых случаях гемоглобин образует патологические соединения. При отравлении угарным газом образуется карбоксигемоглобин (НЬСО). Сродство гемоглобина с окисью углерода значительно выше, чем с кислородом, а скорость диссоциации карбоксигемоглобина в 200 раз меньше, чем оксигемоглобина. Поэтому присутствие в воздухе даже 1% угарного газа приводит к прогрессирующему увеличению количества карбоксигемоглобина и опасному угарному отравлению. Кровь теряет способность переносить кислород. Развивается гипоксия мозга и других тканей. Угарное отравление сопровождается сильной головной болью, тошнотой, рвотой, судорогами, потерей сознания и смертью.
При отравлении сильными окислителями, например нитритами, марганцовокислым калием, красной кровяной солью, образуется метгемоглобин (МеtНЬ). В этом соединении гемоглобина железо становится трехвалентным. Поэтому метгемоглобин очень слабо диссоциирующее соединение. Он не отдает кислород тканям. Все соединения гемоглобина имеют характерный спектр. Восстановленный гемоглобин дает одну широкую полосу поглощения в желто-зеленой части спектра между линиями D и Е. Оксигемоглобина дает 2 узких полосы поглощения в желто-зеленой части спектра между линиями D и Е. У карбоксигемоглобина такая же спектральная картина, как и оксигемоглобина. Поэтому для диагностики отравления угарным газом, в исследуемую кровь добавляют восстановитель, например реактив Стокса. Под их влиянием оксигемоглобин превращается в дезоксягемоглобин и появляется спектр восстановленного гемоглобина. Карбоксигемоглобин не восстанавливается. Метгемоглобин, в зависимости от рН крови, дает 3-5 полос поглощения. Одна из них находится в красной части, другие в желто-зеленой области спектра.
Гемоглобин образует с соляной кислотой соединение коричневого цвета - соляно кислый гематин. Форма его кристаллов зависит от видовой принадлежности крови. В частности, кристаллы соляно кислого гематина человека имеют форму прямоугольных пластинок.
Содержание гемоглобина определяют методом Сали.
Гемометр Сали состоит из 3 пробирок, находящихся в специальном штативе. Две из них, расположенные сбоку от центральной, заполнены стандартным раствором соляно кислого гематина коричневого цвета. Средняя пробирка имеет градуировку в единицах гемоглобина. В нее наливают 0,2 мл соляной кислоты. Затем мерной пипеткой набирают 20 мкл крови и выпускают ее в соляную кислоту. Перемешивают содержимое пробирки и выдерживают 5 мин. Полученный раствор соляно кислого гематина разводят водой до тех пор, пока его цвет не станет таким же. как в боковых пробирках. По уровню жидкости в средней пробирке определяется содержание гемоглобина. В норме в крови мужчин содержится 132-164 г/л (13,2-16.4г.%) гемоглобина. У женщин -115-145 г/л (11,5-14,5 г %). Количество гемоглобина снижается при кровопотерях, интоксикациях, нарушениях эритропоэза. недостатке железа, витамина В12 и т.д.. Кроме этого определяют цветовой показатель. Он отражает степень насыщения эритроцитов гемоглобином. Это отношение содержания гемоглобина в крови к количеству эритроцитов. В норме его величина составляет 0,85-1.05.
Цианметгемоглобиновый фотометрический метод является наиболее точным. Основан на превращении гемоглобина в цианмет-гемоглобин при добавлении к крови определенного количества специального реактива (раствор Драбкина). Показания фотоэлектроколориметра соответствуют определенному содержанию в крови гемоглобина.
47. Гемолиз это разрушение мембраны эритроцитов и выход гемоглобина в плазму. В результате кровь становится прозрачной.
Различают следующие виды гемолиза.
По месту возникновения:
1. Эндогенный, т.е. в организме.
2. Экзогенный, вне его. Например, во флаконе с кровью, аппарате искусственного кровообращения. По характеру:
1. Физиологический. Он обеспечивает разрушение старых и патологических форм эритроцитов. Имеется два механизма. Внутриклеточный гемолиз происходит в макрофагах селезенки, костного мозга, клетках печени. Внутрисосудистый,в мелких сосудах, из которых гемоглобин с помощью белка плазмы гаптоглобина переносится к клеткам печени. Там гем гемоглобина превращается в билирубин. В сутки разрушается около 6-7 г гемоглобина.
2. Патологический.
По механизму возникновения:
I Химический. Возникает при воздействии на эритроциты веществ, растворяющих липиды мембраны. Это спирты, эфир, хлороформ, щелочи, кислоты и т.д. В частности, при отравлении большой дозой уксусной кислоты возникает выраженный гемолиз.
2. Температурный. При низких температурах в эритроцитах образуются кристаллики льда, разрывающие их оболочку.
3. Механический. Наблюдается при механических разрывах мембраны. Например, при встряхивании флакона с кровью или ее перекачивания аппаратом искусственного кровообращения.
4. Биологический. Происходит при действии биологических факторов. Это гемолитические яды бактерий, насекомых, змей. В результате переливания несовместимой крови.
5. Осмотический. Возникает в том случае, если эритроциты попали в среду с осмотическим давлением ниже чем у крови. Вода входит в эритроциты, они набухают и лопаются. Концентрация хлорида натрия, при которой происходит гемолиз 50% всех эритроцитов, является мерой их осмотической стойкости. Ее определяют в клинике для диагностики заболеваний печени, анемии. Осмотическая стойкость должна быть не ниже 0.46% Наd. При помещении эритроцитов в среду, с большим чем у крови осмотическим давлением, происходит плазмолиз. Это сморщивание эритроцитов. Его используют для подсчета эритроцитов.
Скорость оседания эритроцитов (СОЭ).
Оседание эритроцитов — их свойство осаждаться на дне сосуда (капилляр), при сохранении крови в несвертывающемся состоянии в виде так называемых монетных столбиков, над которыми образуется слой прозрачной жидкости — плазмы.
Эритроциты оседают потому, что их относительная плотность больше, чем относительная плотность плазмы.
Факторы, влияющие на СОЭ.
СОЭ зависит от белкового состава плазмы, главным образом от соотношения глобулинов и альбуминов (в норме АГ-коэффициент равен 1,5—2,3).
Клиническое значение.
Сдвиг коэффициента за счет увеличения количества глобулинов ускоряет СОЭ, что имеет место при многих патологических (воспаление, туберкулез) и некоторых физиологических (беременность) процессах.
Все белковые молекулы (фибриноген, а(-глобулин, аг-глобу-лин, (3-глобулин) в разной степени снижают дзета-потенциал эритроцитов — отрицательный заряд мембран, способствующий взаимному отталкиванию эритроцитов и поддержанию их во взвешенном состоянии. Наибольшее влияние оказывают фибриноген, иммуноглобулины, гаптоглобин.
Изменения СОЭ имеют большое диагностическое и прогностическое значение.
48ГРУППЫ КРОВИ. Часто для сохранения жизни при кровопотерях и травмах приходится возмещать кровь. Первое упоминание о переливании крови относится к 1667 г, когда кровь ягненка была перелита человеку. Человек остался жив. Дальнейшие переливания крови оканчивались смертью и французский парламент запретил переливание крови, В 1829 году Блендель (Англия) и в 1832 году Вольтер (Россия) продолжили переливание крови. До 1870 года было проведено 60 переливаний: 16 из них окончились смертью пациента. Почему некоторые переливания оканчивались смертью? Этот вопрос изучали австрийский ученый Карл Ландштейнер и чешский психиатр Ян Янский. При смешивании эритроцитов одних людей с сывороткой других они наблюдали явление изогемагглютинации, т.е. склеивания эритроцитов. Причины склеивания были раскрыты Ландштейнером. В эритроцитах человека были обнаружены агглютиногены - вещества, которые могут быть склеены, т.е. склеиваемые вещества. Различают два вида агглютиногенов, которые обозначают латинскими буквами А и В. Ландштейнер обнаружил, что у людей в эритроцитах могут содержаться агтлютиноген А или В, или же они оба могут отсутствовать. Соответственно этому Ландштейнер и открыл три группы крови. Ян Янский открыл четвертую группу крови, в эритроцитах которой содержатся оба вида агглютиногенов. Янский дал группам крови порядковые обозначения римскими цифрами I, Ц, III, IV. Наряду с агтлютиногенами Ландштейнер в плазме крови обнаружил два вида агглютининов - веществ, которые склеивают, т.е. склеивающие вещества. Они и поныне обозначаются греческими буквами аир. Агглютинины аир являются врожденными антителами. В плазме крови человека может быть агглютинин а или [), могут быть оба вместе или же оба отсутствовать. Эти четыре группы крови в настоящее время принято обозначать также и латинскими буквами.
Группы крови человека передаются по наследству и не меняются.
Агглютинины имеют два центра связывания. Поэтому молекула агглютинина образует мостик между двумя эритроцитами. Значит каждый эритроцит за счет агглютининов связывается с соседними и образуется конгломерат (реакция агглютинации). Реакция агглютинации происходит при встрече агг-лютиногена с одноимённым агглютинином: А с а, В с (J. В плазме наряду с агглютининами имеются гемолизины (их также два вида: о и (3). При встрече агглютиногена с гемолизином происходит гемолиз эритроцитов. Гемолизины действуют при температуре 37-40° С. Поэтому переливание несовместимой крови через 30-40 с приводит к гемолизу эритроцитов. При комнатной температуре происходит только агглютинация, но не наблюдается гемолиз. В плазме крови II, III.IV групп имеются антиагглютинины (во II группе-А, III-В, IV-AB), покинувшие эритроцит и ткани. Они связывают агглютинины и гемолизины крови при переливании небольших ее доз. Но у 10-20% людей имеется высокая концентрация агглютининов и гемолизинов, которые не могут быть связаны с антиагглютининами.
На мембране эритроцитов у людей с первой группой крови находится антиген Н, а у людей с другими группами крови он присутствует на клетках в качестве скрытой детерминанты. Из Н-антигена образуются антигены А, В. Антиген Н у лиц с первой группой крови может быть связан с анти-Н-антителами, которые часто встречаются в крови второй и четвертой групп и реже в третьей. У людей с I группой крови встречаются изоиммунные антитела анти-А и реже анти-В. Значит, наличие антигена-Н и изоиммунных антител может быть причиной гемотрансфузионных осложнений при переливании крови первой группы людям с другими группами. У людей с 1(0) группой крови найдена специфическая субстанция, обозначаемая символом О -это слабый антиген. В настоящее время система АВО часто обозначается АВН, а вместо терминов агглютиноген и агглютинины применяются соответственно антиген и антитела.
С учётом причин агглютинации были сформулированы два основных классических правила переливания крови, которые сводятся к следующему.
1. Чтобы избежать склеивания эритроцитов и последующих осложнений, требуется исключить встречу агглютиногенов донора с одноименными агглютининами реципиента.
2. При переливании крови агглютинины и гемолизины донора в расчет не принимаются, т.к. они разводятся в плазме реципиента, связываются с его антиагглютининами и теряют свою активность, не представляя опасности для эритроцитов.
Из этих схем видно, что эритроциты I (0) группы не склеиваются никакими сыворотками, поэтому их можно вводить всем людям. Реципиентам с четвертой группой крови можно вводить эритроциты людей всех групп крови.
Однако второе классическое правило переливания крови правомерно лишь в тех случаях, когда переливается небольшое количество (200-300 мл) крови. В современной медицинской практике нередко приходится переливать несколько литров крови. Вследствие большого объема переливаемой крови, агглютинины и гемолизины донора в плазме реципиента не разводятся. Наличие Н-антигена и изоиммунных антител у лиц с первой группой крови может приводить к гемотрансфузионным осложнениям при её переливании. Поэтому сейчас при переливании учитывают так называемые современные правила.
Агглютиногены А и В содержатся не только в крови, но и во всех тканях организма. Они устойчивы, переносят нагревание до 100°С и действие различных химических веществ (ацетона, аммиака и т. д.). Поэтому агглюти-ногены А и В обнаруживаются в древних ископаемых останках людей.
49.Резус-фактор
В эритроцитах большинства людей имеется еще один фактор, обнаруженный впервые Ландштейнером и Винером в 1940 г. в крови обезьян макак (macacusresus) и поэтому названный резус-фактором (Rh). Это сильный антиген, который наследуется.
Резус-фактор имеется у 86% людей и они названы резусположительными (Rh+). У 14% людей этот фактор отсутствует, и они названы резусотрицательными (Rh~). Резус-фактор (антиген), хотя и находится в эритроцитах, но не связан с агглютиногенами. В отличие от агглютиногенов у резус-фактора в сыворотке нет агглютининов или антител.
Этот фактор имеет практическое значение потому, что если в кровь резус-отрицательным людям повторно вводить кровь резус-положительных людей, то происходит гемолиз эритроцитов, т. е. в крови резус-отрицательных людей образуются особые антитела против резус-фактора, а реакция между антителом и резус-фактором приводит к повреждению эритроцитов.
Особое значение имеют те случаи, когда резусположительный плод развивается у резус-отрицательной женщины. В этом случае резус-фактор плода диффундирует через плаценту в кровь матери, что приводит к образованию в крови противо-резусных антител. Последние проходят через плаценту в кровь плода и могут вызвать у него тяжелые нарушения вследствие гемолиза эритроцитов. Этим объясняются некоторые случаи мертворождаемости. Однако примерно в 10% случаев гемолитической болезни, хотя и у матери, и у плода кровь резусположи-тельная, анемия все-таки наступает. Было установлено, что резус-фактор бывает трех типов (D, С, Е), и он по типу не совпадает у матери и плода.
Практическое значение при переливании крови имеют 6 антигенов системы Rh-Hr (D, С, Е, d, с, е). Первые три антигена являются разновидностями резус-фактора - D(Rh0), С(rh′ ), Е(rh″ ). Наиболее антигене и является наиболее частой причиной изосерологических конфликтов при гемотрансфузиях и беременности антиген D(Rh0), наиболее слаб - Е(rh″ ). Поэтому при переливании крови необходимо предупредить введение антигена D(Rh0) с кровью донора реципиентам, этот антиген не имеющим. С этой точки зрения резус-принадлежность у реципиентов определяют по наличию антигена D(Rh0), а другие антигены системы Rh-Hr не учитываются.
Если у доноров определять резус-принадлежность по тому же принципу, что и у реципиентов, то оказывается, что в 2-3% случаев резус-отрицательная донорская кровь содержит в эритроцитах антигены С(rh′ ) и Е(rh″ ). В связи с этим к группе доноров с резус-отрицательной кровью должны относиться только лица, в эритроцитах которых нет антигенов D(Rh0), С(rh′ ) и Е(rh″ ). Это обстоятельство имеет существенное значение, так как лицо, в эритроцитах которого обнаружены антигены С(rh′ ) или Е(rh″ ), будучи донором, относится к группе резус-положительной, но, будучи реципиентом, должно считаться резус-отрицательным, ибо отсутствует антиген 0.
Таким образом в крови индивидуума может быть один тип резус-фактора или комбинация из нескольких типов, причем каждый из типов резус-фактора вызывает образование специфических антител.
В эритроцитах есть также антигены системы Hr-Hr0, rh′, rh″, которые вызывают образование специфических антител, однако их антигенные свойства слабее, чем у резус-фактора. Наиболее частой причиной иммунизации оказывается антиген rh′(с), наименее антигенны rh″(е) и Hr0(d). Все лица с резус-отрицательной кровью одновременно являются Hr-положительными, если имеют антиген rh′(с). Наличие антигена Нr заставляет предостеречь от трансфузий резус-отрицательной крови реципиентам с резус-положительной кровью или вообще без определения резус-принадлежности больного, так как можно вызвать иммунизацию или посттрансфузионное осложнение по антигену rh′(с), если больной окажется Hr-отрицательным.
Перед переливанием крови обязательно следует установить резус-принадлежность донора и реципиента и провести пробу на резус-совместимость. При переливании крови следует строго придерживаться принципа использования крови, одноименной по резус-фактору.
Около 80 % людей имеют I и II группы крови, 15 % - III и 5 % - IV группу крови. Отдавать свою кровь для переливания, т. е. быть донором, может каждый здоровый человек. Донорство приносит пользу не только больным, которым переливание крови иногда спасает жизнь, но и самому донору. Взятие небольшого количества крови у человека (200-250 мл) усиливает деятельность кроветворных органов.
50. Лейкоциты. Лейкоциты, или белые кровяные клетки, в свежей крови бесцветны. Число их составляет в среднем 4-9 • 109 л. Лейкоциты в кровяном русле и лимфе способны к активным движениям, могут переходить через стенку сосудов в соединительную ткань органов, где они выполняют основные защитные функции. По морфологическим признакам и биологической роли лейкоциты подразделяют на две группы: зернистые лейкоциты, или гранулоциты, и незернистые лейкоциты, или агранулоциты .
У зернистых лейкоцитов выявляются специфическая зернистость (эозинофильная, базофильная или нейтрофильная) и сегментированные ядра. В соответствии с окраской специфической зернистости различают нейтрофильные, эозинофильные и базофильные гранулоциты.
Функции. Лейкоциты выполняют многообразные функции, направленные прежде всего на защиту организма от агрессивных чужеродных влияний. Одни из них обеспечивают специфический иммунитет, другие осуществляют фагоцитоз микроорганизмов и уничтожение их с помощью ферментов, третьи оказывают бактерицидное действие.
Лейкоциты выполняют также и секреторную функцию: выделяют антитела с антибактериальными и антитоксическими свойствами, ферменты — протеазы, пептидазы, диастазы, липазы и др. За счет этих веществ лейкоциты могут повышать проницаемость капилляров и даже повреждать эндотелий.
Лейкоцитарная формула. Процентное соотношение основных видов лейкоцитов называется лейкоцитарной формулой.
Гранулоциты. К гранулоцитам относятся нейтрофильные, эозинофильные и базофильные лейкоциты. Они образуются в красном костном мозге, содержат специфическую зернистость в цитоплазме и сегментированные ядра.
Нейтрофильные гранулоциты— самая многочисленная группа лейкоцитов, составляющая 2,0—5,5 • 109 л крови. В популяции нейтрофилов крови могут находиться клетки различной степени зрелости — юные, палочкоядерные и сегментоядерные. В цитоплазме нейтрофилов видна зернистость.
Основная функция нейтрофилов — фагоцитоз микроорганизмов, поэтому их называют микрофагами.
Продолжительность жизни нейтрофилов составляет 5—9 сут.
Эозинофильные грамулоциты. Количество эозинофилов в крови составляет 0,02— 0,3 • 109 л.
азурофильные (первичные) и эозинофильные (вторичные).
Функция. Эозинофилы способствуют снижению гистамина в тканях различными путями. Специфическая функция – антипаразитарная.
Базофильные гранулоциты. Количество базофилов в крови составляет 0—0,06 • 109/л
Функции. Базофилы опосредуют воспаление и секретируют эозинофильный хемотаксический фактор, образуют биологически активные метаболиты арахидоновой кислоты — лейкотриены, простагландины.
Продолжительность жизни. Базофилы находятся в крови около 1—2 сут.
Характеристика отдельных видов лейкоцитов
Лейкопоэз осуществляется экстраваскулярно и лейкоциты, в том числе и нейтрофилы, попадают в сосудистое русло благодаря амебовидному движению и выделению протеолитических ферментов, способных растворять белки костного мозга и капилляров. В циркулирующей крови нейтрофилы живут от 8 ч до 7 сут. Находящиеся в кровотоке нейтрофилы могут быть условно разделены на 2 группы: 1) свободно циркулирующие и 2) занимающие краевое положение в сосудах. Между обеими группами существует динамическое равновесие и постоянный обмен. Следовательно, в сосудистом русле нейтрофилов содержится приблизительно в 2 раза больше, чем определяется в вытекающей крови.
Предполагают, что разрушение нейтрофилов происходит за пределами сосудистого русла. По-видимому, все лейкоциты уходят в ткани, где и погибают. Обладая фагоцитарной функцией, нейтрофилы поглощают бактерии и продукты разрушения тканей. В составе нейтрофилов содержатся ферменты, разрушающие бактерии. Нейтрофилы способны адсорбировать антитела и переносить их к очагу воспаления, принимают участие в обеспечении иммунитета.
Под влиянием продуктов, выделяемых нейтрофилами, усиливается митотическая активность клеток, ускоряются процессы репарации, стимулируется гемопоэз и растворение фибринового сгустка.
Базофилы. В крови базофилов очень мало (40—60 в 1 мкл) однако в различных тканях, в том числе сосудистой стенке, содержатся тучные клетки, иначе называемые «тканевые базофилы». Функция базофилов обусловлена наличием в них ряда биологически активных веществ. К ним в первую очередь принадлежит гистамин, расширяющий кровеносные сосуды. В базофилах содержатся противосвертывающее вещество гепарин, а также гиалуроновая кислота, влияющая на проницаемость сосудистой стенки. Кроме того, базофилы содержат фактор активации тромбоцитов — ФАТ (соединение, обладающее чрезвычайно широким спектром действия), тромбоксаны (соединения, способствующие агрегации тромбоцитов), лейкотриены и простагландины — производные арахидоновой кислоты и др. Особо важную роль играют эти клетки при аллергических реакциях, когда под влиянием комплекса антиген — антитело происходит дегрануляция базофилов и биологически активные соединения поступают в кровь, обусловливая клиническую картину перечисленных заболеваний.
Количество базофилов резко возрастает при лейкозах, стрессовых ситуациях и слегка увеличивается при воспалении.
Эозинофилы. Длительность пребывания эозинофилов в кровотоке не превышает нескольких часов, после чего они проникают в ткани, где и разрушаются. Эозинофилы обладают фагоцитарной активно- стью. Особенно интенсивно они фагоцитируют кокки. В тканях эозинофилы скапливаются преимущественно в тех органах, где содержится гистамин — в слизистой оболочке и полслизистой основе желудка и тонкой кишки, в легких. Эозинофилы захватывают гистамин и разрушают его с помощью фермента гистаминазы. В составе эозинофилов находится фактор, тормозящий выделение гистамина тучными клетками и базофилами. Эозинофилы играют важную роль в разрушении токсинов белкового происхождения, чужеродных белков и иммунных комплексов.
. Увеличение числа эозинофилов, наблюдаемое при миграции личинок, является одним из важнейших механизмов в ликвидации гельминтозов.
. При этом эозинофилы выполняют роль «чистильщиков», фагоцитируя и инактивируя продукты, выделяемые базофилами.
В эозинофилах содержатся катионные белки, которые активируют компоненты калликреин-кининовой системы и влияют на свертывание крови.
Моноциты. Циркулируют до 70 ч, а затем мигрируют в ткани, где образуют обширное семейство тканевых макрофагов. Функции их весьма многообразны. Моноциты являются чрезвычайно активными фагоцитами, распознают антиген и переводят его в так называемую иммуногенную форму, образуют биологически активные соединения — монокины (действующие в основном на лимфоциты), играют существенную роль в противоинфекционном и противораковом иммунитете, синтезируют отдельные компоненты системы комплемента, а также факторы, принимающие участие в сосудисто-тромбоцитарном гемостазе, процессе свертывания крови и растворении кровяного сгустка.
Лимфоциты. Как и другие виды лейкоцитов, образуются в костном мозге, а затем поступают в сосудистое русло. Здесь одна популяция лимфоцитов направляется в вилочковую железу, где превращается в так называемые Т-лимфоциты (от слова thymus).
Популяция Т-лимфоцитов гетерогенна и представлена следующими классами клеток. Т-киллеры, или убийцы (от англ. tu kill — убивать), осуществляющие лизис клеток-мишеней, к которым можно отнести возбудителей инфекционных болезней, грибки, микобактерии, опухолевые клетки и др. Т-хелперы, или помощники иммунитета. Различают Т—Т-хелперы, усиливающие клеточный иммунитет, и Т—В-хелперы, облегчающие течение гуморального иммунитета. Т-амплифайеры усиливают функцию Т- и В-лимфоцитов, однако в большей степени влияют на Т-лимфоциты. Т-супрессоры — лимфоциты, препятствующие иммунному ответу. Различают Т—Т-супрессоры, подавляющие клеточный иммунитет, и Т—В-супрессоры, угнетающие гуморальный иммунитет. Т-дифференцирующие, или Td-лимфоциты, регулируют функцию стволовых кроветворных клеток, т. е. влияют на соотношение эритроцитарного, лейкоцитарного и тромбоцитарного (мегакариоцитарного) ростков костного мозга. Т-контрсупрессоры препятствуют действию Т-супрессоров и, следовательно, усиливают иммунный ответ. Т-клетки памяти хранят информацию о ранее действующих антигенах и таким образом регулируют так называемый вторичный иммунный ответ, который проявляется в более короткие сроки, так как минует основные стадии этого процесса.
Другая популяция лимфоцитов образует В-лимфоциты (от слова bursa), окончательное формирование которых у человека и млекопитающих, по-видимому, происходит в костном мозге или системе лимфоидно-эпителиальных образований, расположенных по ходу тонкой кишки (лимфоидные, или пейеровы бляшки и др.).
Большинство В-лимфоцитов в ответ на действие антигенов и цитокинов переходит в плазматические клетки, вырабатывающие антитела и потому именуемые антителопродуцентами. Среди В-лимфоцитов также различают В-киллеры, В-хелперы и В-супрессоры.
В-киллеры выполняют те же функции, что и Т-киллеры. Что касается В-хелперов, то они способны представлять антиген, усиливать действие Td-лимфоцитов и Т-супрессоров, а также участвовать в других реакциях клеточного и гуморального иммунитета. Функция В-cynpeccopoв заключается в торможении пролиферации антителопродуцентов, к которым принадлежит основная масса В-лимфоцитов.
Существует группа клеток, получивших наименование «ни Т-, ни В-лимфоциты». К ним относятся так называемые 0-лимфоциты, являющиеся предшественниками Т- и В-клеток и составляющие их резерв. Большинство исследователей относят к 0-лим-фоцитам особые клетки, именуемые натуральными (природными) киллерами, или НК-лимфоцитами. Как и другие цитотоксические лимфоциты (ЦТЛ), НК-лимфоциты секретируют белки, способные «пробуравливать» отверстия (поры) в мембране чужеродных клеток и потому названные перфоринами. ЦТЛ содержат протеолитические ферменты (цитолизины), которые проникают в чужеродную клетку через образующиеся поры и разрушают ее.
Существуют клетки, несущие на своей поверхности маркеры Т- и В-лимфоцитов (двойные клетки). Они способны заменять как те, так и другие.
51 Лейкоцитарная формула
В норме и патологии учитывается не только количество лейкоцитов, но и их процентное соотношение, получившее наименование лейкоцитарной формулы, или лейкограммы
В крови здорового человека могут встречаться зрелые и юные формы лейкоцитов, однако в норме обнаружить их удается лишь у самой многочисленной группы — нейтрофилов. К ним относятся юные и палочкоядерные нейтрофилы. Юные нейтрофилы, или миелоциты, имеют довольно крупное бобовидное ядро, палочкоядерные — содержат ядро, не разделенное на отдельные сегменты. Зрелые, или сегментоядерные, нейтрофилы имеют ядро, разделенное на 2 или 3 сегмента. Чем больше сегментов в ядре, тем старее нейтрофил. Увеличение количества юных и палочкоядерных нейтрофилов свидетельствует об омоложении крови и носит название сдвига лейкоцитарной формулы влево, снижение количества этих клеток свидетельствует о старении крови и называется сдвигом лейкоцитарной формулы вправо. Сдвиг влево часто наблюдается при лейкозах (белокровие), инфекционных и воспалительных заболеваниях.
52.Остановка кровотечения, т.е. гемостаз может осуществляться двумя путями. При повреждении мелких сосудов она происходит за счет первичного или сосудисто-тромбоцитарного гемостаза. Он обусловлен сужением сосудов и закупоркой отверстия склеившимися тромбоцитами. При повреждении этих сосудов происходит прилипание или адгезия тромбоцитов к краям раны. Из тромбоцитов начинают выделяться АДФ, адреналин и серотонин. Серотонин и адреналин суживают сосуд. Затем 'АДФ вызывает агрегацию, т.е. склеивание тромбоцитов. Это обратимая агрегация. После, под влиянием тромбина, образующегося в процессе вторичного гемостаза, развивается необратимая агрегация большого количества тромбоцитов. Образуется тромбоцитарный тромб, который уплотняется, т.е. происходит его ретракция. За счет первичного гемостаза кровотечение останавливается в течение 1-3 минут. Вторичный гемостаз или гемокоагуляция, это ферментативный процесс образования желеобразного сгустка - тромба. Он происходит в результате перехода растворенного в плазме белка фибриногена в нерастворимый фибрин. Образование фибрина осуществляется в несколько этапов и при участии ряда факторов свертывания крови. Они называются прокоагулянтами, так как до кровотечения находятся в неактивной форме. В зависимости от местонахождения факторы свертывания делятся на плазменные, тромбоцитарные, тканевые, зритроцитарные и лейкоцитарные. Основную роль в механизмах тромбообразования играют плазменные и тромбоцитарные факторы.
Выделяют следующие плазменные факторы, обозначаемые римскими цифрами:
1. Фибриноген. Это растворимый белок плазмы крови.
II. Протромбин, оксиглобулин.
III. Тромбопластин. Комплекс фосфолипидов, выделяющийся из тканей и тромбоцитов при их повреждении.
IV. Ионы кальция.
V. Проакцелерин В-глобулин.
VI. Изъят из классификации, так как является активным V фактором.
VII. Проконвертин В-глобулин.
VIII. Антигемофильный глобулин А. В-глобулин.
IX. Антигемофильный глобулин В. Фактор Кристмаса. Фермент протеаза.
X. Фактор Стюарта-Прауэра.
XI. Плазменный предшественник тромбопластина. Фактор Розенталя. Иногда называют антигемофильным
глобулином.
XII. Фактор Хагемана. Протеаза.
XIII. Фибрин-стабилизирующий фактор. Транспептидаза. Все плазменные про коагулянты, кроме тромбопластина и ионов кальция синтезируются в печени. Имеется 12 тромбоцитарных факторов свертывания. Они обозначаются арабскими цифрами. Основные из них:
3. Участвует в образовании плазменной протромбиназы.
4. Антагонист гепарина.
6. Тромбостенин. Вызывает укорочение нитей фибрина.
10. Серотонин. Суживает сосуды, ускоряет свертывание крови.
Выделяют три фазы свертывания крови.
I. Образование активной протромбиназы. Существует 2 ее формы - тканевая и плазменная. Тканевая образуется при выделении поврежденными тканями тромбопластина и его взаимодействии с IV, V, VII и Х плазменными прокоагулянтами. Тромбопластин и VII фактор проконвертин, активируют Х фактор - Стюарта-Прауэра. После этого Х фактор связывается с V - проакцелерином. Этот комплекс является тканевой протромбиназой. Для этих процессов нужны ионы кальция. Это внешний механизм активации процесса свертывания. Его длительность 15 сек.
Внутренний механизм запускается при разрушении тромбоцитов. Он обеспечивает образование плазменной протромбиназы. В этом процессе участвуют Тромбопластин тромбоцитов, IV, V, VIII, IX, X, XI и XII плазменные факторы и 3 тромбоцитарный. Тромбопластин активирует XII фактор Хагемана, который вместе с 3 фактором
тромбоцитов переводит в активную форму XI, фактор Розенталя. Активный XI фактор активирует IX -антигемофильный глобулин В.. После этого формируется комплекс из активного IX фактора, VIII - антигемофильного глобулина А, 3 тромбоцитарного фактора и ионов кальция. Этот комплекс обеспечивает активацию Х фактора - Стюарта-Прауэра. Комплекс активного X, V фактора - проакцелерина и 3 фактора тромбоцитов является плазменной протромбиназой. Продолжительность этого процесса 2-10 мин.
II. Переход протомбина в тромбин. Под влиянием протромбиназы и IV фактора — ионов кальция, протромбин
переходит в тромбин. В эту же фазу под действием тромбина происходит необратимая агрегация тромбоцитов.
III. Образование фибрина. Под влиянием тромбина, ионов кальция и XIII –фибрин-стабилизирующего фактора,
фибриноген переходит в фибрин. На первом этапе под действием тромбина фибриноген, расщепляется на 4 цепи фибрина, мономера. Соединяясь между собой, они формируют волокна фибрина-полимера. После этого XIII фактор, активируемый ионами кальция и тромбином, стимулирует, образование: прочной сети нитей, фибрина. В этой сети задерживаются форменные элементы крови. Возникает тромб. На этом процессе тромбообразования не заканчивается. Под влиянием б фактора тромбоцитов-тромбостенина нити фибрина укорачиваются. Происходит ретракция т.е. уплотнение тромба. Одновременно сокращающиеся нити фибрина стягивают края раны, что способствует ее заживлению.
При отсутствии какого-либо прокоагулянта свертывание крови нарушается. Так как жирорастворимый витамин К имеет исключительное значение для синтеза протомбина, VII, [X и Х плазменных факторов, его недостаток в печени ведет к нарушению механизмов свертывания. Это наблюдается при нарушениях функций печени, ухудшении всасывания жиров, угнетении желчеобразования.
53Механизмы регуляции кроветворения
Кровь и органы, в которых происходят образование и разрушение клеток крови, а также органы, участвующие в перераспределении крови, составляют систему крови. К ней относятся: кровь, циркулирующая в сосудах, красный костный мозг, селезенка, лимфатические узлы и печень.
В красном костном мозге из единой так называемой стволовой клетки возникают все виды клеток крови. Различают три ростка крови: 1) лейкоцитарный, из которого возникают гранулоциты, моноциты и лимфоциты; 2) эритроцитарный, продуцирующий эритроциты; 3) мегакариоцитарный, из которого образуются тромбоциты.
Стволовые недифференцированные клетки, по внешнему виду похожие на лимфоцит, способны к самоподдержанию и к дифференцировке по всем рядам клеток крови. Дифференцировка стволовой полипотентной клетки представляет собой многостадийный процесс, начинающийся с образования морфологически сходных между собой клеток-предшественниц каждого ростка. Затем происходит пролиферация и постепенная дифференцировка клеток, специфическая для каждого ряда, в виде гранулоцитопоэза (трех видов гранулоцитов), моноцитопоэза, лимфоцитопоэза (В- и Т-лимфоцитов), эритроцитопоэза. В период развития клетки приобретают морфологические, гистохимические и функциональные особенности каждого вида форменных элементов крови и на определенной стадии теряют способность к размножению.
Считают, что дифференцировку клеток крови определяют клетки стромы органа, в котором происходит эта дифференцировка. Костные мозговые стромальные клетки определяют дифференцировку в сторону миелоцитопоэза, т. е. процессов грануломоноцитопоэза, эритроцитопоэза и тромбоцитопоэза. Клетки селезенки определяют дифференцировку в сторону лимфопоэза.
Важным фактором в регуляции кроветворения оказалась поэтиночувствительность клеток — предшественниц всех трех ростков. Принципиальное значение этого свойства выражается в том, что поэтиночувствительные клетки способны отвечать на дистантные гуморальные раздражения. Гемопоэтины стимулируют пролиферацию кроветворных клеток. Различают эритропоэтины, лейкопоэтины и тромбоцитопоэтины.
Регуляция эритропоэза. В норме у человека в крови поддерживается постоянное число эритроцитов. В сутки образуется примерно 200—250 млрд. эритроцитов. В то же время ежедневно 0,8% эритроцитов разрушается и столько же продуцируется вновь. Если происходит повышенное разрушение эритроцитов, то их образование усиливается. Стимулятором эритропоэза является эритропоэтин, Основным местом образования эритропоэтина являются почки. Возможно, что эритропоэтин вырабатывается в некотором количестве в других органах и тканях. Наиболее частым стимулом образования эритропоэтина является гипоксия, которая вызывает усиленное размножение кроветворных клеток. В частности, повышенное образование эритропоэтина и эритроцитов происходит в высокогорной местности с низким парциальным давлением кислорода в воздухе, при кровопотерях, при массовом разрушении эритроцитов, при гемолитической анемии.
Для образования нормального количества эритроцитов нужны и другие гуморальные стимуляторы. К активным факторам кроветворения относятся витамин B12 и фолиевая кислота. Первый в 1000 раз активнее, чем фолиевая кислота. Витамин В12 назван внешним фактором кроветворения, так как поступает в организм с пищей. Он содержится в мясе, яйцах, дрожжах. Витамин B12 может всасываться лишь в том случае, если в желудке имеется так называемый внутренний фактор кроветворения, который представляет собой мукопротеид, выделяемый железами желудка. Витамин B12 взаимодействует с внутренним фактором, образуя биологически активный комплекс, который всасывается, откладывается в печени, оттуда поступает в костный мозг, где стимулирует эритропоэз. Эритропоэз ослабляется, если снижается поступление витамина B12 извне или уменьшается выработка внутреннего фактора, обеспечивающего всасывание данного витамина. В этих случаях развивается заболевание крови — В12-дефицитная анемия. Существенное влияние на кроветворение могут оказать белки плазмы, гормоны, особенно гипофиза, и продукты обмена веществ.
Эритроциты разрушаются в селезенке, печени и костном мозге. Чем больше эритроцитов разрушается, тем больше образуется продуктов распада, стимулирующих кроветворение; тем самым обеспечивается приспособительное саморегулирование кроветворения.
Регуляция лейкопоэза. Срок жизни различных видов лейкоцитов варьирует от нескольких часов до нескольких недель. Лейкоциты разрушаются в ретикулярной ткани и на поверхности слизистой оболочки пищеварительного тракта. Наряду с разрушением беспрерывно образуются новые лейкоциты. На количество лейкоцитов в крови влияют гуморальные и нервные факторы.
Лейкопоэз стимулируется продуктами распада самих лейкоцитов. Чем больше разрушается лейкоцитов, тем больше их образуется. Стимулирующее действие на лейкопоэз оказывают продукты распада тканей при их повреждении или при развитии воспалительного процесса. Введение в организм белков или нуклеиновых кислот приводит к нейтрофильному лейкоцитозу с появлением молодых форм нейтрофилов (палочкоядерных, миелоцитов). Аналогичным образом действуют пирогены (вещества, повышающие температуру тела), эндотоксины бактерий, гормоны коркового слоя надпочечников (глюкокортикои-Ды), гипофиза (адренокортикотропный гормон, соматотропный гормон). Повышение количества лейкоцитов в этих случаях развивается в течение нескольких часов и обусловлено выходом нейтрофилов из костномозгового гранулоцитарного резерва. Вирусная инфекция и антигены ведут к усиленной продукции и повышению количества лимфоцитов в крови. Предполагается,. что воздействия, повышающие образование лейкоцитов, приводят к выработке лейкопоэтинов, хотя химический состав и место образования последних еще не выяснены.
Нервная система оказывает определенное влияние на содержание лейкоцитов в крови. Возбуждение симпатической нервной системы, болевое раздражение, эмоциональное возбуждение ведут к увеличению количества лейкоцитов в крови. Повышается их содержание в крови при усиленной мышечной работе и во время пищеварения вследствие перераспределительного лейкоцитоза. Он происходит за счет выброса в общий круг кровообращения лейкоцитов, находящихся в депо крови — в селезенке и в синусах костного мозга. На пищеварительный перераспределительный лейкоцитоз можно выработать условный рефлекс.
Органы системы крови содержат большое количество рецепторных аппаратов. В экспериментальных условиях показано, что раздражение этих рецепторов вызывает различные рефлекторные реакции. Существует двусторонняя связь между органами системы крови и центральной нервной системой. Органы системы крови получают импульсы из центральной нервной системы, которые регулируют их состояние. Вместе с тем органы системы крови являются источником рефлексов, изменяющих состояние как их самих, так и других систем организма. Эти механизмы направлены . на поддержание постоянства состава крови, как внутренней среды организма.
Регуляция тромбоцитопоэза. Это явление менее всего изучено. Обнаружено, что фильтраты плазмы здоровых людей обладают тромбоцитопоэтической активностью. Тромбоцитопоэтины связаны с высокомолекулярной белковой фракцией, относящейся к у-глобулинам. Тромбоцитопоэтины неоднородны. Одни из них стимулируют образование тромбоцитов в костном мозге, другие— выход тромбоцитов в кровь.
54. Сердечнососудистая система - физиологическая система, включающая сердце, кровеносные сосуды, лимфатические сосуды, лимфатические узлы, лимфу, механизмы регуляции (местные механизмы: периферические нервы и нервные центры, в частности сосудодвигательный центр и центр регуляции деятельности сердца).
Таким образом, сердечно-сосудистая система - это совокупность 2-х подсистем: системы кровообращения и системы лимфообращения. Сердце - основной компонент обеих подсистем.
Кровь и лимфа беспрерывно движутся по сосудам человеческого тела, которые густой сетью оплетают его органы и ткани. В рез-те такого движения крови и лимфы осуществляется доставка кислорода и питательных веществ клеткам, удаление продуктов обмена веществ и гуморальная регуляция деятельности органов и систем в организме.
По хар-ру циркулирующей жидкости сосудистую систему человека и позвоночных можно разделить на два отдела: кровеносный — систему сосудов, по которым циркулирует кровь (артерии, капилляры, вены и сердце) и лимфатический — систему сосудов, по которым движется лимфа. Лимфатическая система, вступая в процессе эмбриогенеза в связь с кровеносной системой, является добавочным для венозных сосудов руслом. Движение жидкости в лимфатических сосудах происходит так же, как и в венах, в направлении от тканей к центру.
Кровеносная система состоит из центрального органа —сердца и находящихся в соединении с ним замкнутых трубок различного калибра, называемых кровеносными сосудами. Сердце ритмическими сокращениями приводит в движение всю массу крови, содержащуюся в сосудах, которые тоже способствуют кровообращению. Кровеносные сосуды, идущие от сердца и несущие кровь к органам, называются артериями. Артерии, постепенно разветвляясь и уменьшая диаметр, переходят в арте-риолы, а артериолы —в мельчайшие сосуды — капилляры, по которым артериальная кровь подходит непосредственно к тканям. Здесь происходит обмен газов между кровью и тканями, кровь превращается в венозную и переходит в венулы. Венулы, сливаясь, образуют сначала мелкие, а затем крупные вены, по которым кровь возвращается к сердцу.
МАЛЫЙ И БОЛЬШОЙ КРУГ КРОВООБРАЩЕНИЯ
Кровообращение начинается в тканях, где совершается обмен веществ и газов через стенки капилляров (кровеносных и лимфатических, ).Капилляры составляют основную часть микроциркуляторного сосудистого русла. Микрогемоциркуляторное русло включает следующие пять звеньев: артерио-лы, прекапиллярные артериолы, являющиеся промежуточным звеном между артериолами и истинными капиллярами, капилляры, посткапиллярные венулы и венулы. Из микрогемоциркуляторного русла кровь поступает в вены, а лимфа — в лимфатические сосуды, которые в конечном счете впадают в присер-дечные вены. Венозная кровь, содержащая присоединившуюся к ней лимфу, вливается в полость сердца, сначала в правое предсердие, а из него в правый желудочек. Из последнего венозная кровь поступает в легкие по малому кругу кровообращения.
Малый (легочный) круг кровообращения служит для обогащения крови кислородом в легких. Он начинается в правом желудочке сердца, куда переходит через правое предсердно-желудочковое отверстие венозная кровь. Из правого желудочка выходит легочный ствол, который разветвляется в легких на артерии, переходящие в капилляры. В капиллярных сетях, оплетающих легочные пузырьки (альвеолы), кровь отдает углекислоту и получает кислород, при этом кровь снова приобретает алый цвет и превращается в артериальную. Обогащенная кислородом артериальная кровь поступает из капилляров в вены, которые, слившись в четыре легочные вены (по две с каждой стороны), впадают в левое предсердие сердца. В левом предсердии заканчивается малый (легочный) круг кровообращения, а поступившая в предсердие артериальная кровь переходит через левое предсердно-желудочковое отверстие в левый желудочек, где начинается большой круг кровообращения.
Большой (телесный) круг кровообращения служит для доставки всем органам и тканям кислорода и питательных веществ. Он начинается в левом желудочке сердца,. из которого выходит аорта, несущая артериальную кровь. Артериальная кровь содержит необходимые для жизнедеятельности организма питательные вещества и кислород. Цвет ее ярко-алый. Аорта разветвляется на артерии, которые идут ко всем органам и тканям тела и переходят в толще их в артериолы и далее в капилляры. Капилляры собираются в венулы и затем — в вену. Через стенку капилляров происходит обмен веществ и газообмен между кровью и тканями тела. Протекающая в капиллярах артериальная кровь отдает в ткани питательные вещества и кислород и взамен получает продукты обмена и углекислоту. Поступающая в венозное русло кровь бедна кислородом и богата углекислотой, поэтому венозная кровь имеет темную окраску. При кровотечении по цвету крови можно определить, какой сосуд поврежден — артерия или вена. Вены сливаются в два крупных ствола — верхнюю и нижнюю полые вены, которые впадают в правое предсердие сердца. Этим отделом сердца заканчивается большой круг кровообращения.
Дополнением к большому кругу является третий круг кровообращения, обслуживающий само сердце. Он начинается выходящими из аорты венечными артериями сердца и заканчивается венами сердца. Последние сливаются в венечный синус, впадающий в правое предсердие, а остальные вены открываются в полость предсердия непосредственно.
Самые крупные из артерий: 2 сонные, питающие головной мозг. Коронарные –питают сердце, подключичные снабжают кровью верхние конечности, почечные и подвздошные- нижние конечности.
Магистральные сосуды(аорта, крупные артерии) – непрерывность кровотока
Сосуды сопротивления( артериоллы, прекапилляры) – оптимальный уровень кровяного давления, имеют толстые стенки поэтому от диаметра зависит сколько крови поступает в органы
Шунтирующие (артерио-венозные анастомозы)- обеспечивают переход крови из артериол в венулы минуя капилляры.
Обменные сосуды, или истинные капилляры, — обмен между кровью и тканями.
Емкостные сосуды- венозные сосуды, крупные, стенка тонкая
Резистивные сосуды концевые артерии, артериоллы, прекапилляры
Сосуды дыпо- селезенка, печень, верхушка легких, подкожные сплетения
Благодаря постоянному движению крови в сосудах, обеспечиваются основные функции системы кровообращения:
1) транспорт веществ, необходимых для обеспечения функций клеток организма;
2) доставка к клеткам организма химических веществ, регулирующих их обмен;
3) отвод от клеток, переработанных в них, веществ (метаболитов);
4) гуморальная, т.е. осуществляемая через жидкость, связь органов и тканей между собой;
5) доставка тканям средств защиты;
6) удаление вредных веществ из организма;
7) обмен тепла в организме.
55Сердце человека располагается в грудной клетке. Это четырехкамерный мышечный орган, бессменно работающий в течение всей жизни. По форме сердце напоминает уплощенный конус и состоит из двух частей — правой и левой. Каждая часть включает предсердие и желудочек. Величина сердца приблизительно соответствует величине кулака человека. Масса сердца в среднем около 300 г. У тренированных к мышечной работе людей размеры сердца больше, чем у нетренированных. Сердце покрыто тонкой и плотной оболочкой, образующей замкнутый мешок — околосердечную сумку. Между сердцем и околосердечной сумкой находится жидкость, увлажняющая сердце и уменьшающая трение при его сокращениях. Мышечная стенка желудочков значительно толще стенки предсердий. Это объясняется тем, что желудочки выполняют большую работу по перекачиванию крови по сравнению с предсердиями. Особенной толщиной отличается мышечная стенка левого желудочка, который, сокращаясь, проталкивает кровь по сосудам большого круга кровообращения. Предсердия и желудочки соединяются между собой отверстиями. По краям отверстий располагаются створчатые клапаны сердца. На стороне клапанов, обращенной в полость желудочков, имеются специальные сухожильные нити. Эти нити
удерживают клапаны от прогибания. Между левым предсердием и левым
желудочком клапан имеет две створки и называется двустворчатым, между правым предсердием и правым желудочком находится трехстворчатый клапан. Двустворчатый и трехстворчатый клапаны обеспечивают ток крови в одном направлении — из предсердий в желудочки. Между левым желудочком и отходящей от него аортой, а также между правым желудочком и отходящей от него легочной артерией тоже имеются клапаны. Из-за своеобразной формы створок они названы полулунными. Каждый полулунный клапан состоит из трех листков, напоминающих кармашки. Свободным краем кармашки направлены в просвет сосудов. Полулунные клапаны обеспечивают ток крови только в одном направлении — из желудочков в аорту и легочную артерию. Предсердия и желудочки могут находиться в двух состояниях: сокращенном
и расслабленном. Сокращение и расслабление предсердий и желудочков сердца происходят в определенной последовательности и строго согласованы времени. Сердечный цикл состоит из сокращения предсердий, сокращения желудочков, расслабления желудочков и предсердий (общего расслабления).
Стенка сердца состоит из 3х слоёв.
Эндокард - тонкий соединительнотканный слой переходящий в tunica intima сосудов. Обеспечивает несмачиваемость стенки сердца, облегчая внутрисосудистую гемодинамику.
Миокард - миокард предсердия отделяется от миокарда желудочков фиброзным кольцом.
Эпикард - состоит из 2-х слоёв - фиброзный (наружный) и сердечный (внутренний). Фиброзный листок окружает сердце снаружи - выполняет защитную функцию и предохраняет сердце от растяжения.
Сердечный листок состоит из 2-х частей:
висцеральный (эпикард);
париетальный, который срастается с фиброзным листком.
Между висцеральным и париетальным листками есть полость, заполненная жидкостью (уменьшает травмы).
Значение перикарда:
защита от механических повреждений;
защита от перерастяжения.
Оптимальный уровень сердечного сокращения достигается при увеличении длинны мышечных волокон не более чем на 30-40 % от исходной величины. Обеспечивает оптимальный уровень работы клеток синсатриального узла. При перерастяжении сердца нарушается процесс генерации нервных импульсов. Опора для крупных сосудов (препятствует спадению полых вен).
56Правильное направление обеспечивает клапанный аппарат сердца:
трехстворчатый;
легочный;
митральный;
аортальный клапаны
Они открываются в нужный момент и закрываются, препятствуя кровотоку в обратном направлении.
Трехстворчатый клапан
Расположен между правым предсердием и правым желудочком. Он состоит из трех створок. Если клапан открыт, кровь переходит из правого предсердия в правый желудочек. Когда желудочек наполняется, мышца его сокращается и под действием давления крови клапан закрывается, препятствуя обратному току крови в предсердие.
Легочный клапан
При закрытом трехстворчатом клапане выход крови в правом желудочке возможен только через легочной ствол в легочные артерии. Легочный клапан расположен на входе в легочный ствол. Он открывается под давлением крови при сокращении правого желудочка, кровь поступает в легочные артерии, затем под действием обратного тока крови при расслаблении правого желудочка он закрывается, препятствуя обратному поступлению крови из легочного ствола в правый желудочек.
Двустворчатый или митральный клапан
Находится между левым предсердием и левым желудочком. Состоит из двух створок. Если он открыт, кровь поступает из левого предсердия в левый желудочек, при сокращении левого желудочка он закрывается, препятствуя обратному току крови.
Аортальный клапан
Закрывает вход в аорту. Тоже состоит из трех створок, которые имеют вид полулуний. Открывается при сокращении левого желудочка. При этом кровь поступает в аорту. При расслаблении левого желудочка, закрывается. Таким образом, венозная кровь (бедная кислородом) из верхней и нижней полой вен попадает в правое предсердие. При сокращении правого предсердия через трехстворчатый клапан она продвигается в правый желудочек. Сокращаясь, правый желудочек выбрасывает кровь через легочной клапан в легочные артерии (малый круг кровообращения). Обогащаясь кислородом в легких кровь превращается в артериальную и по легочным венам продвигается в левое предсердие, затем в левый желудочек. При сокращении левого желудочка артериальная кровь через аортальный клапан под большим давлением попадает в аорту и разносится по всему организму (большой круг кровообращения).
При работе сердца возникают звуки, которые называют тонами сердца. При выслушивании (аускультации) тонов сердца на поверхности левой половины грудной клетки слышны два тона: I тон (систолический), II тон — в начале диастолы (диастолический). Тон I более протяжный и низкий, II — короткий и высокий.
Детальный анализ тонов сердца стал возможным благодаря применению электронной аппаратуры. Если к груди обследуемого приложить чувствительный микрофон, соединенный с усилителем и осциллографом, можно зарегистрировать тоны сердца в виде кривых — фонокардиограммы (ФКГ). Эта методика называется фонокардиографией
Сужение клапанных отверстий или неплотное смыкание створок и лепестков клапанов вызывает появление сердечных шумов, возникающих вследствие вихреобразного (турбулентного) движения крови через отверстия клапанов. Эти шумы имеют важное диагностическое значение при поражениях клапанов сердца.
На ФКГ, помимо I и II тонов, регистрируются III и IV тоны сердца (более тихие, чем I и II, поэтому неслышные при обычной аускультации).
Тон III возникает вследствие вибрации стенки желудочков при быстром притоке крови в желудочки в начале их наполнения.
Тон IV имеет два компонента. Первый из них возникает при сокращении миокарда предсердий, а второй появляется в самом начале расслабления предсердий и падения давления в них.
К внешним проявлениям деятельности сердца относят артериальный пульс, характер которого отражает не только деятельность сердца, но и функциональные состояния артериальной системы. Артериальный пульс отражает ритм сердца, скорость изгнания крови левым желудочком и величину систолического объема, т. е. факторы, определяющие кинетическую энергию выброшенной сердцем крови. Это в какой-то мере позволяет судить о силе сердечных сокращений.
Эхокардиография основана на использовании ультразвука для получения изображения сердца и крупных сосудов. Ультразвуковые волны частотой 18-20 кГц, превышающей разрешающую способность человеческого уха, распространяются как продольные колебания со скоростью, которая зависит от физических свойств вещества, через которое они проходят. Они генерируются пьезоэлектрическими кристаллами под воздействием переменного электрического поля, которые осуществляют преобразование электрической и механической (звуковые колебания) энергии друг в друга и функционируют одновременно как передатчик звука и приемник отраженных звуковых волн (эхо-волн). Получение изображения структур сердца с помощью ультразвука основано на отражении ультразвуковых волн на границе между двумя веществами с разными физическими свойствами, как, например, кровью и эндокардом. Поскольку при этом угол падения равен углу отражения, получаемое изображение является зеркальным.
Эхокардиография позволяет увидеть расположение и характер движения различных структур сердца (клапанов, стенок полостей и др.), и поэтому является важнейшим методом изучения многих аспектов анатомии и физиологии сердца. Ее достоинствами являются неинвазивный характер, относительная дешевизна, возможность применения у постели больного, быстрота получения специфичной, количественной, воспроизводимой и надежной информации, благодаря чему эхокардиография приближается к идеальному методу диагностики.
Основное ограничение эхокардиографии связано с элементами субъективности при получении и интерпретации эхо-изображения, что предъявляет высокие требования к квалификации исследователя. Из-за отчасти эмпирического подбора интенсивности сигнала отдельные структуры могут не визуализироваться и, наоборот, могут появляться артефакты. Не является 100 % и воспроизводимость результатов измерений и величин расчетных показателей. Диагностическое значение эхокардиографии в М-режиме и двухмерной эхокардиографии:
1) верификация диагноза: выпотного перикардита; стеноза митрального (и трехстворчатого) клапанов, обструктивной формы гипертрофической кардиомиопатии (идиопатического гипертрофического субаортального стеноза); опухоли предсердия; пролапса митрального клапана;
2) получение важных данных для диагностики: врожденных и приобретенных пороков сердца; инфекционного эндокардита; тромбов в полостях сердца; констриктивного перикардита; ИБС; застойной сердечной недостаточности; кардиомиопатии; легочной гипертензии; аневризмы аорты, в том числе расслаивающей;
3) точное измерение размеров структур сердца: поперечного размера левого желудочка и левого предсердия; толщины межжелудочковой перегородки и стенок левого желудочка; диаметра корня аорты;
4) анализ движения структур сердца: клапанов; межжелудочковой перегородки; свободных стенок левого и правого желудочков;
57.Весь сердечный цикл длится 0,8-0,86 с.
Две основные фазы сердечного цикла:
систола - выброс крови из полостей сердца в результате сокращения;
диастола - расслабление отдых и питание миокарда, наполнение полостей кровью.
Эти основные фазы подразделяются на:
систола предсердий - 0,1 с - кровь поступает в желудочки;
диастола предсердий - 0,7 с;
систола желудочков - 0,3 с - кровь поступает в аорту и лёгочный ствол;
диастола желудочков - 0,5 с;
общая пауза сердца - 0,4 с. Желудочки и предсердия в диастоле. Сердце отдыхает, питается, предсердия наполняются кровью и на 2/3 напонляются желудочки.
Сердечный цикл начинается в систоле предсердия. Систола желудочка начинается одновременное диастолой предсердий.
Цикл работы желудочков (Шово и Морели (1861 г.)) - состоит из систолы и диастолы желудочков.
Систола желудочков: период сокращения и период изгнания.
Сердечный цикл состоит из сокращения предсердий и желудочков и последующего расслабления. У здорового человека в состоянии покоя сердце сокращается 60-70 раз в 1 мин.
Во время общего расслабления сердца (диастола) кровь из полых и легочных вен поступает соответственно в правое и левое предсердия. После этого наступает сокращение (систола) предсердий, процесс сокращения начинается у места впадения верхней полой вены в правое предсердие и распространяется
по обоим предсердиям, в результате чего кровь из предсердий через предсердно-желудочковые отверстия нагнетается в желудочки. Затем в стенках сердца начинается волна сокращения (систола) желудочков, которая распространяется на оба желудочка, и кровь из них нагнетается в отверстия легочного ствола и аорты, в это время предсердно-желудочковые клапаны закрываются. После этого наступает пауза. Возвращению крови из аорты и легочного ствола в желудочки препятствуют полулунные клапаны. Описанное последовательное сокращение и расслабление различных отделов сердца связано с проводящей системой.
Началом каждого цикла считается систола предсердий, длящаяся 0,1 сек. В это время миокард желудочков расслаблен, створчатые клапаны открыты, а полулунные закрыты. Во время сокращения предсердий вся кровь из них поступает в желудочки. По окончании систолы предсердий начинается систола желудочков, которая длится 0,3 сек. Во время сокращения желудочков предсердия оказываются уже расслабленными, двухстворчатый и трехстворчатый клапаны закрываются. При сокращении миокарда желудочков кровь из них выталкивается в аорту и легочный ствол. Полулунные клапаны у начала аорты и легочного ствола открываются, их заслонки прижимаются изнутри к стенкам этих сосудов, и кровь из желудочков направляется в аорту и легочный ствол.
Сокращение желудочков сменяется их расслаблением - диастолой желудочков. Под действием высокого давления, создающегося в аорте и легочном стволе, их полулунные клапаны закрываются, препятствуя возвращению крови в желудочки. После этого наступает период покоя всех камер сердца, или общего расслабления, - общая пауза, которая длится 0,4 сек. Такого интервала между сокращениями достаточно для полного восстановления работоспособности сердца. Во время сокращения (систолы) желудочков кардиомиоциты не способны отвечать на новые раздражения. Этот периодневозбудимости миокарда называется рефрактерной фазой. Свойство сердечной мышцы во время систолы не отвечать дополнительным сокращением позволяет миокарду более полно и ритмично сокращаться, не испытывая утомления.
В связи с изменением давления в полостях сердца клапаны сердца, легочной артерии и аорты открываются или закрываются. В начале систолы желудочков предсердно-желудочковые клапаны закрываются, а полулунные клапаны аорты и легочной артерии открываются. В период диастолы желудочков происходит систола предсердий, предсердно-желудочковые клапаны открываются и желудочки заполняются кровью. Возвращению крови из аорты и легочного ствола препятствуют полулунные клапаны.
58Сердечная мышца, как и скелетная, обладает возбудимостью, способностью проводить возбуждение и сократимостью.
Возбудимость сердечной мышцы. Сердечная мышца менее возбудима, чем скелетная. Для возникновения возбуждения в сердечной мышце необходимо применить более сильный раздражитель, чем для скелетной. Установлено, что величина реакции сердечной мышцы не зависит от силы наносимых раздражений (электрических, механических, химических и т. д.). Сердечная мышца максимально сокращается и на пороговое, и на более сильное по величине раздражение.
Проводимость. Волны возбуждения проводятся по волокнам сердечной мышцы и так называемой специальной ткани сердца с неодинаковой скоростью. Возбуждение по волокнам мышц предсердий распространяется со скоростью 0,8—1,0 м/с, по волокнам мышц желудочков— 0,8—0,9 м/с, по специальной ткани сердца—2,0—4,2 м/с.
Сократимость. Сократимость сердечной мышцы имеет свои особенности. Первыми сокращаются мышцы предсердии, затем—папиллярные мышцы и субэндокардиальный слой мышц желудочков. В дальнейшем сокращение охватывает и внутренний слой желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.
Физиологическими особенностями сердечной мышцы является удлиненный рефрактерный период и автоматия. Теперь о них поподробнее.
Рефрактерный период. В сердце в отличие от других возбудимых тканей имеется значительно выраженный и удлиненный рефрактерный период. Он характеризуется резким снижением возбудимости ткани в течение ее активности. Выделяют абсолютный и относительный рефрактерный период (р.п.). Во время абсолютного р.п. какой бы силы не наносили раздражения на сердечную мышцу, она не отвечает на него возбуждением и сокращением. Он соответствует по времени систоле и началу диастолы предсердий и желудочков. Во время относительного р.п. возбудимость сердечной мышцы постепенно возвращается к исходному уровню. В этот период мышца может ответить на раздражитель сильнее порогового. Он обнаруживается во время диастолы предсердий и желудочков.
Сокращение миокарда продолжается около 0.3 с, по времени примерно совпадает с рефрактерной фазой. Следовательно, в период сокращения сердце неспособно реагировать на раздражители. Благодаря выраженному р.п. .р.рррр.п., который длится больше чем период систолы, сердечная мышца неспособна к тетаническому (длительному) сокращению и совершает свою работу по типу одиночного мышечного сокращения.
Автоматия сердца. Вне организма при определенных условиях сердце способно сокращаться и расслабляться, сохраняя правильный ритм. Следовательно, причина сокращений изолированного сердца лежит в нем самом. Способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом, носит название автоматии.
В сердце различают рабочую мускулатуру, представленную поперечнополосатой мышцей, и атипическую, или специальную, ткань, в которой возникает и проводится возбуждение.
У человека атипическая ткань состоит из:
синоатриального узла, располагающегося на задней стенке правого предсердия у места впадения полых вен;
атриовентрикулярного (предсердно-желудочкого) узла находящегося в правом предсердии вблизи перегородки между предсердиями и желудочками;
пучка Гиса (председно-желудочковый пучок), отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через перегородку между предсердиями и желудочками, делится на две ножки, идущие к правому и левому желудочкам. Заканчивается пучок Гиса в толще мышц волокнами Пуркинье. Пучок Гиса—это единственный мышечный мостик, соединяющий предсердия с желудочками.
Синоаурикулярный узел является ведущим в деятельности сердца (водитель ритма), в нем возникают импульсы, определяющие частоту сокращений сердца. В норме атриовентрикулярный узел и пучок Гиса являются только передатчиками возбуждения из ведущего узла к сердечной мышце. Однако им присуща способность к автоматии, только выражена она в меньшей степени, чем у синоаурикулярного узла, и проявляется лишь в условиях патологии.
Атипическая ткань состоит из малодифференцированных мышечных волокон. В области синоаурикулярного узла обнаружено значительное количество нервных клеток, нервных волокон и их окончаний, которые здесь образуют нервную сеть. К узлам атипической ткани подходят нервные волокна от блуждающих и симпатических нервов.
В отличии от скелетных мышц сердечная мышцы работает только при наличии кислорода. Сердце проходит только аэробный процесс окисления. А в скелетных анаэробный еще может. Скелетные мышцы питаются глюкозой. Сердечная мышца питается ненасыщенными жирными кислотами70%, глюкозой 30%,