

Лекция 2
Структурно-функциональные особенности эритроцитов.
Гемоглобин. СОЭ
Раздел I. Эритроциты
Ключевая функция
Эритроциты - это форменные элементы крови, осуществляющие дыхательную функцию, – перенос кислорода от легких к тканям и двуокиси углерода от тканей к легким. Отсутствие ядра не позволяет определять эритроцит как клетку крови, но в научной литературе термин «клетка» в приложении к эритроциту используется.
Форма
Эритроциты имеют форму двояковогнутого диска (рис. 1). Диаметр эритроцитов варьирует в пределах от 7,6 мкм до 8 мкм, наибольшая толщина составляет 2,4 мкм.
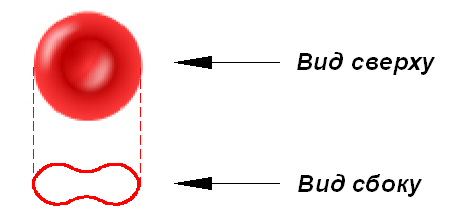
Рис. 1. Эритроцит, вид сверху и сбоку
Вогнутость мембраны позволяет оптимально насытить эритроциты кислородом за короткое время прохождения клеток крови по капиллярам малого круга.
 Уникальная
форма эритроцита
Уникальная
форма эритроцита
во-первых, увеличивает его поверхность: чем больше поверхность клетки, тем больше молекул кислорода, диффундирующих в плазму крови из легких, растворяется в мембране эритроцита и поступает в его цитоплазму.
|
во-вторых, уменьшает время диффузии кислорода до молекул гемоглобина, удаленных от плазматической мембраны. |
Заряд мембраны
Наружная поверхность мембраны эритроцитов имеет выраженный отрицательный заряд. Заряд эритроцита обусловлен химическим строением сиаловых кислот, содержащих СОО-- группы (рис. 2).
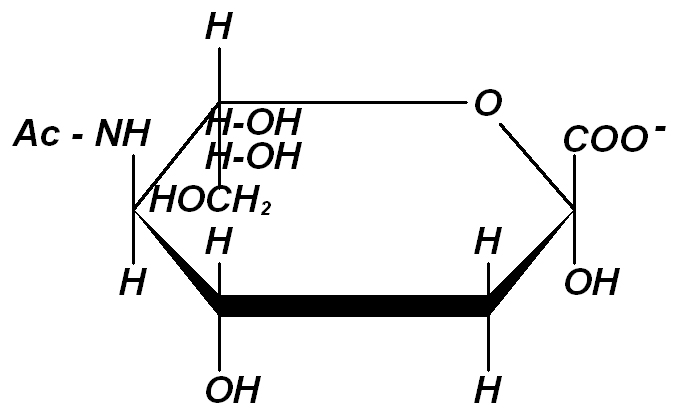
Рис. 2. Строение одной из сиаловых кислот -
N-aцетилнейраминовой кислоты
Cиаловые кислоты являются частью гликокаликса – олигосахаридных структур, связанных с наружной поверхностью мембраны клетки. Эти кислоты соединяются с N-концевым фрагментом интегрального белка мембраны эритроцитов гликофорина (рис. 3).
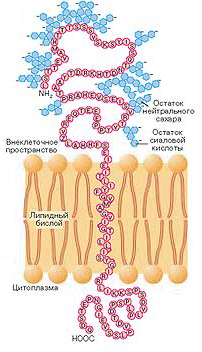
Рис. 3. Структура гликофорина
Отрицательный заряд мембраны исключает возможность сближения эритроцитов друг с другом и с отрицательно заряженной стенкой сосуда. На поверхности стареющих эритроцитов количество сиаловых кислот снижается, что увеличивает вероятность образования «монетных столбиков» из сблизившихся эритроцитов.
Проницаемость мембраны.
Na+/К+-насос
 Мембрана
эритроцита избирательно проницаема:
Мембрана
эритроцита избирательно проницаема:
 хорошо
плохо
хорошо
плохо
для анионов (СI- , НСО3- ) для катионов (Na+, К+)
Cвободное перемещение анионов через мембрану из плазмы крови в клетку и наоборот связано с участием эритроцитов в сохранении рН крови. Полагают, что в переносе анионов через мембрану участвуют некоторые интегральные белки мембраны эритроцитов.
Катионы, несмотря на низкую проницаемость, также стремятся диффундировать через мембрану вдоль градиента концентрации, поскольку ионов Na+ в плазме крови значительно больше, чем в цитоплазме эритроцита, а количество ионов К+, напротив, преобладает в эритроците. Содержание Na+ и К+, однако, поддерживается в пределах нормальных значений постоянно действующей ферментативной системой мембраны эритроцита, осуществляющей активный перенос ионов против их концентрационных градиентов – Na+/К+-насосом.
Деформируемость эритроцита
Эритроциты обладают обратимой деформируемостью. Это свойство позволяет переносчикам кислорода проходить через капилляры, диаметр которых меньше диаметра эритроцитов, и восстанавливать исходную форму, покидая узкие сосуды. Способность эритроцитов изменять продольный и поперечный размеры в основном обеспечивается структурной организацией ключевого белка цитоскелета эритроцита – спектрина.
Молекулы спектрина имеют фибриллярную конформацию. Соединяясь друг с другом, они образуют упругую сеть на некотором расстоянии от внутренней поверхности мембраны эритроцита. Благодаря химическим связям с рядом протеинов мембраны, спектрин взаимодействует с интегральными белками мембраны. Эти связи делают сопряженными изменения конформации цитоскелета и интегральных белков мембраны.
В крупных сосудах эритроциты пребывают в постоянном вращательном движении. В капиллярах клетки красной крови перестают вращаться, существенно удлиняются, их поверхность возрастает. При этом происходит перемешивание молекул гемоглобина в цитоплазме эритроцита. Эти факторы увеличивают эффективность насыщения эритроцита кислородом.