

Структура і склад біомембран
Мембрани складаються з ліпідів трьох класів: фосфоліпіди, гліколіпіди і холестерол. Фосфоліпіди і гліколіпіди (ліпіди з приєднаними до них вуглеводами) складаються з двох довгих гідрофобних вуглеводневих "хвостів", які пов'язані із зарядженою гідрофільній "головою". Холестерол надає мембрані жорсткість, займаючи вільний простір між гідрофобними хвостами ліпідів і не дозволяючи їм згинатися. Тому мембрани з малим вмістом холестеролу більш гнучкі, а з більшим - більш жорсткі й тендітні. Також холестерол служить "стопором", що перешкоджає переміщенню полярних молекул з клітки і в клітку. Важливу частину мембрани складають білки, які пронизують її і відповідають за різноманітні властивості мембран. Їх склад і орієнтація в різних мембранах розрізняються.Клітинні мембрани часто асиметричні, тобто шари відрізняються за складом ліпідів, перехід окремої молекули з одного шару в інший (так званий фліп-флоп) утруднений.
Мембранні органели
Це замкнуті одиночні або пов'язані один з одним ділянки цитоплазми, відділені від гіалоплазми мембранами. До одномембранним органел відносяться ендоплазматична мережа, апарат Гольджі, лізосоми, вакуолі, пероксисоми; до двумембранним - ядро, мітохондрії, пластиди. Будова мембран різних органел відрізняється за складом ліпідів і мембранних білків.
Виборча проникність
Клітинні мембрани мають виборчої проникністю: через них повільно дифундують глюкоза, амінокислоти, жирні кислоти, гліцерин і іони, причому самі мембрани певною мірою активно регулюють цей процес - одні речовини пропускають, а інші ні. Існує чотири основних механізми для надходження речовин у клітину або виведення їх з клітини назовні: дифузія, осмос, активний транспорт і екзо-або ендоцитоз. Два перших процесу носять пасивний характер, тобто не потребують витрат енергії; два останні - активні процеси, пов'язані зі споживанням енергії. Виборча проникність мембрани при пасивному транспорті обумовлена спеціальними каналами - інтегральними білками. Вони пронизують мембрану наскрізь, утворюючи свого роду прохід. Для елементів K, Na і Cl є свої канали. Щодо градієнта концентрації молекули цих елементів рухаються в клітину і з неї. При подразненні канали натрієвих іонів розкриваються, і відбувається різке надходження в клітку іонів натрію. При цьому відбувається дисбаланс мембранного потенціалу. Після чого мембранний потенціал відновлюється. Канали калію завжди відкриті, через них в клітку повільно потрапляють іони калію.ендоплазматичний ретикулум.Ендоплазматичний ретикулум (ЕПР, від лат. reticulum — «сіточка») або ендоплазматична мережа — внутрішньоклітинна органела еукаріотичних клітин, що представляє собою розгалужену систему з оточенихмембраною сплющених порожнин, бульбашок і канальців.
Будова
Схематичне представлення клітинного ядра, ендоплазматичного ретикулума і комплексу Гольджі.
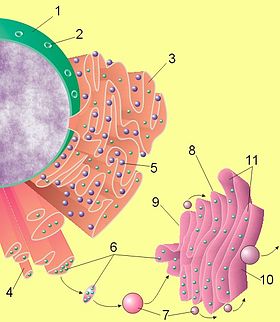
Рис.4
Ядро клітини.
Пори ядерної мембрани.
Шорсткий (гранулярний) ендоплазматичний ретикулум.
Гладкий (агранулярний) ендоплазматичний ретикулум.
Рибосоми на поверхні щорсткого ендоплазматичного ретикулума.
Білки, що транспортуються.
Транспортні везикули.
Комплекс Гольджі.
Ендоплазматичний ретикулум складається з розгалуженої мережі трубочок і кишень, оточених мембраною. Площа мембран ендоплазматичного ретикулума складає більше половини загальної площі всіх мембран клітини.
Мембрана ЕПР морфологічно ідентична оболонці клітинного ядра і складає з нею одне ціле. Таким чином, порожнини ендоплазматичного ретикулума відкриваються в міжмембранну порожнину ядерної оболонки. Мембрани ЕПС забезпечують активний транспорт ряду елементів проти градієнту концентрації. Бульбашки та канальці, що створюють ендоплазматичний ретикулум, мають в поперечнику 0,05-0,1 мікрона (іноді до 0,3 мікрона), товщина двошарових мембран, що стінку канальців, становить близько 50 ангстрем. Ці структури містять ненасичені фосфоліпіди, а також деяка кількість холестерину і сфінголіпідів. У їхній склад також входять білки. Найтонші трубочки, діаметр яких коливається в межах 1000-3000 ангстрем, заповнені гомогенним вмістом та зполучають більші за розміром частини ендоплазматичного ретикулума.
Ендоплазматичний ретикулум не є стабільною структурою і схильний до частих змін. Виділяють два типи ЕПР:
Шорсткий (гранулярний) ендоплазматичний ретикулум,
Гладкий (агранулярний) ендоплазматичний ретикулум.
На поверхні шорсткого ендоплазматичного ретикулума знаходиться велика кількість рибосом, які відсутні на поверхні гладкого ЕПР.
Шорсткий та гладкий ендоплазматичний ретикулум виконують деякі різні функції в клітині.
Функції ендоплазматичного ретикулум
За участю ендоплазматичного ретикулума відбувається трансляція і транспорт мембранних білків, що сектеруються, синтез і транспорт ліпідів і стероїдів. Для ЕПС характерний також накопичення продуктів синтезу. Ендоплазматичний ретикулум бере участь у тому числі і в створенні нової ядерної оболонки (наприклад післямітозу). Ендоплазматичний ретикулум містить внутріклітинний запас кальцію, який є медіатором багатьох реакцій відповіді клітини, зокрема скорочення м'язових клітин. У клітинах м'язових волокон розташована особлива форма ендоплазматичного ретикулума — саркоплазматичний ретикулум.
Функції гладкого ендоплазматичного ретикулума
Гладкий ендоплазматичний ретикулум бере участь в багатьох процесах метаболізму. Ферменти гладкого ендоплазматичного ретикулума беруть участь в синтезі ліпідіві фосфоліпідів, жирних кислот і стероїдів. Також агранулярний ендоплазматичний ретикулум грає важливу роль у вуглеводному обміні, знезараженні клітки і запасанні кальцію. Зокрема, у зв'язку з цим в клітках надниркових залоз і печінки переважає гладкий ендоплазматичний ретикулум.
Синтез гормонів.
До гормонів, які утворюються в гладкому ЕПС, належать, наприклад, статеві гормони хребетних тварин і стероїдні гормони (що синтезуються в надниркових залозах). Клітини яєчок і яєчників, що відповідають за синтез гормонів, містять велику кількість гладкого ЕПР.
Накопичення і перетворення вуглеводів.
Вуглеводи в організмі накопичуються в печінці у вигляді глікогену. За допомогою гліколізу глікоген в печінці трансформується в глюкозу, що є найважливішим процесом в підтримці рівня глюкози в крові. Один з ферментів гладкого ЕПС відщеплює від першого продукту гліколізу, глюкоза-6-фосфату, фосфогрупу, дозволяючи, таким чином, глюкозі залишити клітину і підвищити рівень цукру в крові.
Нейтралізація отрут.
Гладенький ендоплазматичний ретикулум клітин печінки бере активну участь в нейтралізації токсинів. Ферменти гладенького ЕПР приєднують до нерозчинних радикалів гідрофільні послідовності, внаслідок чого токсин стає краще розчинним і може виводитися з клітини, а згодом і з організму через сечу. У разі безперервного надходження отрут, медикаментів або алкоголю, площа гладенького ЕПР збільшується, що дозволяє нейтралізувати більшу кількість токсину за рахунок збільшення робочої поверхні ЕПР.
Саркоплазматичний ретикулум.
Особлива форма гладкого ендоплазматичного ретикулума, саркоплазматичний ретикулум, утворюється в м'язових клітинах, де іони кальцію активно закачуються зцитоплазми в порожнині ЕПР проти градієнту концентрації в не збудженому стані клітини і звільняються в цитоплазму для ініціації скорочення. Концентрація іонів кальцію в ЕПС може досягати 10−3 моль/л, тоді як в цитоплазмі — порядка 10−7 моль/л (в стані спокою). Таким чином, мембрана саркоплазматичного ретикулума забезпечує активне перенесення проти градієнтів концентрації великих порядків. І прийом і звільнення іонів кальцію в ЕПС знаходиться в жорсткій залежності від фізіологічних умов
Концентрація іонів кальцію в цитозолі впливає на безліч внутрішньоклітинних і міжклітинних процесів, таких як активація або інгібування ферментів, експресія генів,синаптична пластичність нейронів, скорочення м'язових клітин, звільнення антитіл з клітин імунной системи.
Функції шорсткого ендоплазматичного ретикулума.
Шорсткий ендоплазматичний ретикулум має дві функції: синтез білків і виробництво мембран.Синтез білків. Білки, що виробляються клітиною, синтезуються в рибосомах, які можуть бути приєднані до поверхні ЕПС, саме це і надає поверхні «шорсткий» вигляд[2]. На поверхні ЕПР синтезується багато білків, призначених для сортування і транслокації до певних відділів клітини, тобто інтегральні мембранні білки, білки органел і білки, призначені для секреції. Проте, рибосоми на поверхні ЕПР на завжди знаходяться там, а приєднуються до нього після початку синтезу білка, призначеного для сортування в ЕПР[3]. Вільна рибосома синтезує білок, поки цитозольні частинки розпізнавання сигналу на впізнають сигнал з 5-15 гідрофобних амінокислот (N-сигнальна послідовність) після позитивно зярядженої амінокислоти на початку ланцюжка. Цей сигнальний фрагмент залишається в мембрані до кінця синтезу, після чого він відрізається від готового білка.
Отримані поліпептідні ланцюги поміщаються в порожнини шорсткого ендоплазматичного ретикулума, де згодом піддаються першим крокам посттрансляційної модифікації і згортаються. Таким чином, лінійні ланцюги амінокислот отримують після транслокації в ендоплазматичний ретикулум необхідну тривимірну структуру.Синтез мембран[ред.]За допомогою виробництва фосфоліпідів, ЕПР розширює також поверхню своїх власних мембран, які за допомогою транспортних везикул транспортуються до інших частин мембранної системи
3.Цитоплазма
Загальна характеристика цитоплазми. Цитоплазма (від грец.kytos — вмістилище, тут — клітина і plasma — виліплене, оформлене) — основний вміст живої клітини, що являє собою гетерогенний комплекс гіалоплазми з розміщеними у ній органоїдами та включеннями. Зовні Ц. оточена плазма лемою, а від вакуоль відмежована тонопластом. В молодих клітинах цитоплазма займає весь простір клітини , а з віком починає структуруватись біля стінок у вигляді тяжів. Під світловим мікроскопом цитоплазма — гомогенна структура. В якій виділяються пластиди та мітохондрії. Субмікроскопічна структура цитоплазми. За допомогою світлового мікроскопа була встановлена субмікроскопічна будова цитоплазми. Було виявлено, що цитоплазма складається з гомогенної рідкої маси гіалоплазми (матрикс цитоплазми ) і складної системи мембран: плазмолеми, тонопласт та ін. Гіалоплазма (від грец. hyalos — скло і plasma — виліплене) — основна оптично прозора частина цитоплазми рослинних клітин, в якій містяться ядро, всі органоїди та продукти внутріклітинного метаболізму. Вона являє собою колоїдну фазу клітини певної в’язкості і здатної до активного руху, бере участь у внутрішньоклітинному транспорті речовин. Вона має складний хімічний склад. Містить проміжні продукти обміну, амінокислоти, РНК, ліпіди, ряд ферментів, необхідних для синтезу білків, нуклеїнових кислот, жирних кислот та ін. сполук. В гіалоплазмі виробляються агрегати макромолекул білка — мікро трубочки, або мікрофіламенти ( від грец. micros — малий від лат. filamentum — нитка)— надмолекулярні агрегати еукаріотичних клітин, які складаються із білкових субодиниць, що формують спіралізовану стрічку. М. беруть участь у зміні клітини при амебоїдному русі, ендомітозі, переміщенні прикріплених до них органел, генерують рух гіалоплазми тощо. Це нестійкі структури білкових компонентів діаметром біля 2,5 нм. , завдовжки кілька мікрометрів. Вони виконують особливі функції і в процесі поділу клітини — беруть участь в переміщені хромосом, у підтриманні форми про пластид, в орієнтації мікрофібрил клітинної оболонки. Мікро трубочки виявлені у всіх типах рослинних клітин. Деякі автори описують їх як окремі органели. Функції гіалоплазми:
Є основною магістраллю для переміщення метаболітів клітини.
Заповнює частину простору плазмодесмових каналів і таким чином забезпечує міжклітинні зв’язки;
Вступаючи в безпосередні контакти з мембранами органел, вона регулює фізико-хімічні і ферментні зв’язки між ними. Структура біологічних мембран цитоплазми. Основні біологічні мембрани цитоплазми. Одним з великих відкриттів електронної мікроскопії — є встановлення мембранної організації цитоплазми. Біологічні мембрани — обов’язкові і дуже важливі її компоненти. Біологічні мембрани представляють собою щільні, дуже тонкі плівки товщиною від 4 до 10 нм. Їх важлива властивість — напівпроникнення. Це пояснює те, що одні речовини, розчинені у воді проходять через мембрани легко навіть проти градієнта концентрації, для інших же речовин мембрани є важко проникним бар’єром. Можна сказати, що мембрани контролюють хімічний склад цитоплазми в цілому і її органел зокрема. Загальна маса мембран в активно функціонуючих клітинах становить 90 % всієї сухої речовини цитоплазми. Структура і хімічний склад мембран неоднорідний, що пов’язано з їх функцією. Структурну основу мембран становлять молекули ліпідів, які формують подвійний бімолекулярний шар. Причому, молекули фосфоліпідів орієнтовані гідрофільними полюсами назовні у напрямок водного середовища (тобто в цитоплазму), а гідрофобними залишками вони орієнтовані всередину. Тобто, вони володіють амфіпатичними властивостями: один кінець молекули гідрофобний, другий — гідрофільний. Гідрофільні кінці полярні, оскільки несуть електричний заряд, гідрофобні кінці не полярні. Молекули білків розташовуються розсіяно по обидва боки ліпідної основи. Частина білкових молекул глибоко занурена у ліпідний каркас, інколи вони розривають його, утворюючи гідрофільні пори. Різноманітність розташування білкових молекул або їх агрегатів в мембранах зумовлюють неоднорідність мембран і їх високу пластичність (лабільність) — здатність змінювати структуру відповідно за зміною функції. Найважливіше значення з біологічних мембран мають два пограничні шари: плазмалеми і тонопласт. Плазмалема (від грец. plasma — виліплене, оформлене і lemma — оболонка) — тонка цитоплазматична мембрана, що межує з клітинною оболонкою. Плазмалему можна побачити під електронним мікроскопом при збільшенні 50 і більше раз. Товщина 7,5- 9,5 нм. П. Має хвилясту, складчасту поверхню, завдяки чому збільшується поверхня всмоктування і безпосередньо прилягає до стінки клітини. Функції плазмалеми:
П. регулює обмін речовин клітини з навколишнім середовищем (вибіркова здатність до проникнення речовин);
Виконує деякі синтетичні функції, зокрема на ній відбувається утворення целюлозних мікрофібрил клітинної оболонки, відіграє велику роль у явищах піноцитозу, фагоцитозу тощо.
Тонопласт (від грец. tonos — напруження і plastos — виліплений, сформований) — біологічна мембрана клітин, що відокремлює цитоплазму від вакуолі і володіє вибірковою проникністю та здатністю до активного транспорту іонів. Т. Ще називають вакуолярною мембраною. Біологічні та фізичні властивості цитоплазми. Цитоплазмі властиві ознаки, які визначають поняття “життя”: рух, ріст, живлення, дихання, подразливість та ін. Цитоплазма — “необхідний субстрат життя” для всіх живих компонентів рослинної клітини. Важливі з них такі: біосинтез білків (рибосоми), фотосинтез (хлоропласти), дисиміляція (мітохондрії), виділення (апарат Гольджі). Зупинимось на одній з біологічних властивостей — русі. Рух цитоплазми — переміщення цитоплазми з усіма її органоїдами відносно оболонки клітини. Спостерігається в клітинах одноклітинних та багатоклітинних організмів. Основними причинами, що зумовлюють рух цитоплазми є постійні обмінні процеси, а головними умовами є наявність тепла і кисню. Рухаючись, цитоплазма захоплює за собою органоїди, тому про її рух можна судити, спостерігаючи за рухом органоїдів. Швидкість руху цитоплазми в різних клітинах неоднакова і в основному залежить від внутрішніх та зовнішніх факторів. Не з однаковою швидкістю рухається маса цитоплазми в межах однієї клітини. Так, шар цитоплазми, що прилягає до оболонки рухається найповільніше. Рух цитоплазми можна стимулювати підвищенням температури, освітленням, а також внутрішніми збудниками, зокрема етиловим спиртом. Встановлено, що для елодеї канадської оптимальна температура для руху цитоплазми є 37˚С. Існує два типи руху цитоплазми:
Коловий;
Струмочковий .
Коловий рух цитоплазми спостерігається в клітинах, у яких протопласт зосереджений біля оболонки клітини, а внутрішня частина клітини виповнена великою вакуолею. Даний тип руху може відбуватися за і проти годинникової стрілки. Швидкість руху цитоплазми не велика. Так, наприклад, в елодеї становить 10-15 м. на секунду, а у валіснерії — 10-20 м. на секунду. Струмочковий рух цитоплазми відбувається в клітинах, у яких цитоплазма розташовується у вигляді тяжів, які перетинають центральну вакуолю. У таких випадках цитоплазма рухається в різних напрямках у виглідя тонких струмочків. Не менш важливою біологічною ознакою цитоплазми є її обмін речовин з навколишнім середовищем. Обмін відбувається при наявності двох протилежних процесів — асиміляції (анаболізму) та дисиміляції (катаболізму). Анаболізм (асиміляція) (від грец. anabole — накидаю, підйом ) — сукупність хімічних процесів, що протікають в організмі і внаслідок яких утворюються складові елементи клітини і тканин з накопиченням енергії. Прикладом анаболізму, що має планетарне значення є фотосинтез. Катаболізм (дисиміляція) (від грец. dissimilis — несхожий) —процес розщеплення в живих клітинах органічних сполук на простіші речовини з вивільненням енергії. Д. взаємозв’язана з протилежним процесом в обміні речовин — асиміляцією. За фізичним станом цитоплазма є колоїдним розчином, тобто таким в якому розчинені речовини не розпадаються до молекул або йонів, а залишаються у підвішеному стані, або представляють собою гігантські макромолекули. При необхідності колоїди цитоплазми віддають воду і переходять у стан геля (стан, при якому колоїдні частинки втрачають заряди). Таке явище спостерігається в насінні, яке знаходиться в стані спокою. Нормальний стан рівноваги цитоплазми, коли її колоїди несуть електричні заряди називається станом золя. Цей стан спостерігається в клітинах насіння, яке починає проростати. Завдяки такій зміні стану цитоплазми насіння в стані спокою може переносити температуру –80-90˚С, тоді як в проростках насіння цитоплазма дегенерує при –50-60˚С. Колоїдний стан цитоплазми в різних клітинах тіла рослин неоднаковий, тому що колоїди знаходяться в різних дисперсних фазах. Неоднакова також стійкість колоїдів цитоплазми рослин із різних географічних широт, що обумовлено історичним розвитком рослин. Так, наприклад, сосна, що зростає в лясах Японії переносить морози до –50-60˚С, тоді як деякі рослини Закавказзя (гранати, інжир, евкаліпти) побиваються морозами –15˚С і гинуть. В інституті низьких температур гілка берези перенесла критичну температуру –278˚С і почала вегетувати і утворила листки. Колоїдний стан цитоплазми може порушуватися під впливом умов навколишнього середовища, а також механічних пошкоджень, що пов’язано з втратою електричних зарядів колоїдними частинками і порушення гідратації оболонок. Це явище називається коагуляцією. У стані коагуляції клітини гинуть.
Хімічний склад цитоплазми. Хімічний склад цитоплазми непостійний і змінюється під впливом реакцій, які в ній протікають. Вміст води коливається від 70-90%, білка — від 10-20%, ліпідів— 2-3%, вуглеводів— 1-2%, мінеральних солей — 1%. Реакція цитоплазми близька до нейтральної. Основною речовиною цитоплазми є білки: прості (протеїни) і складні (протеїди). Білки — це складні високомолекулярні органічні речовини. Елементарний склад білків не постійний. Вони в основному складаються з вуглецю, водню, кисню, азоту та сірки. Макромолекули білків дуже складні. Білки становлять біля 70% сухої ваги цитоплазми. Висока пластичність білків, здатність їх до швидких перебудов обумовлюють значну роль, яку вони відіграють в житті живої клітини. Білки здатні з’єднуватись з іншими компонентами цитоплазми небілкової природи, утворюючи постійні сполуки протеїди: ліпопротеїди (сполуки ліпоїдів з білками), глюкопротеїди (сполуки вуглеводів з білками), нуклеопротеїди (сполуки нуклеїнових кислот з білками), фосфопротеїди (сполуки фосфорної кислоти з білками. До складних білкових компонентів належать також ферменти — біологічні каталізатори, які найчастіше складаються з двох компонентів — білкової частини та небілкової, приєднаної до неї (йон металу, який має змінну валентність Cu, Mg, Fe). Жири (ліпоїди) утворюють велику групу різноманітних сполук. До складу жирів входять вуглець, водень, кисень. Ліпоїди нерозчинні у воді але розчиняються в органічних розчинниках (бензині, ефірі і т.д.). до ліпоїдів належать прості жири або масла, воски, фосфоліпоїди, глюколіпоїди. До цієї ж групи належать каротиноїди і хлорофіл — розчинні в жирах пігменти. Вуглеводи в живій клітині відіграють роль запасних поживних речовин, використовуються як енергетичний матеріал. Вони складаються з вуглецю, водню і кисню. Поділяються на: моно-, ди- та полісахариди. Моносахариди, або прості цукри мають загальну формулу (C6 H12 O6 ) . до цієї групи належать представники: глюкоза та фруктоза. Дисахариди мають складнішу формулу (C12 H22 O11 ) . найбільш поширеним представником є буряковий цукор. Полісахариди мають формулу (C6 H10 O5 ) . До цієї групи належить крохмаль, інулін, геміцелюлоза. Крохмаль — це один з найбільш поширених запасних речовин. Розрізняють первинний крохмаль, який синтезується в хлоропластах та вторинний, який синтезується в лейкопластах — амілопластах. Таким чином, з наведеної характеристики органічних сполук видно, що хімічний склад цитоплазми дуже складний і постійно змінюється.