

2. Сложность алгоритмов и программ, сложность дифференциации
1. Пусть алгебраическая, дифференцирующая активность клеточного генома приводиться в действие и сопровождается процессом, состоящим из двух сортов элементарных актов: запускающих актов в количестве lsi и исполнительных в количестве msi . Причем, их величины таковы, что
![]() (1)
(1)
Язык
из lsi
и msi
команд, удовлетворяющих (1) называется,
согласно [7, Л-1] экономным языком, поскольку
при использовании lsi
- количества «слов» как элементов
мно-жества запускающих команд можно
составить не более, чем
![]() подмножеств -«предложений» или инструкций.
Отметим, что уже в этом определении
автор [7, Л-1] по сути дела рас-сматривал
иерархию команд, во главе которой стоит,
как оказалось, гомеобокс с аналогичной
управляющей «ролью», который был открыт
в 1983-1984 гг.
подмножеств -«предложений» или инструкций.
Отметим, что уже в этом определении
автор [7, Л-1] по сути дела рас-сматривал
иерархию команд, во главе которой стоит,
как оказалось, гомеобокс с аналогичной
управляющей «ролью», который был открыт
в 1983-1984 гг.
2.
Пусть число всех видов актов xsi,
используемых (s,i)-
классом клеток от момента времени нуль
(например, от оплодотворения) до момента
t0=s![]() равно числу всех подмножеств множества,
элементы которого состоят только из
запускающих актов, введенных в действие
на этом временном интервале.
равно числу всех подмножеств множества,
элементы которого состоят только из
запускающих актов, введенных в действие
на этом временном интервале.
3. Число клеток nsi в (s,i) - классе, продифференцировавшем на интервале [0, tsi ] частично или полностью, обратно пропорционально числу элементарных актов хsj : nsi~1/ xs,s+1-i , j = i -1.
При этом предполагалось, что время «изготовления» клетки прямо пропорционально числу всех «строитель-ных» операций, пропорционально ее сложности. Отсюда следует, что полное число однотипных клеток - обратно пропорционально ее сложности. При этом не исключа-ется и «запараллеленные» процессы дифференцировки, т.к. в хsi могут входить целые алгоритмические «блоки в соответствующем исчислении.»
4. Число клеток bsi , продиффренцировавшихся хотя бы частично за время равно
![]() (2)
(2)
Последнее равенство позволило в [7, л-1] постулировать

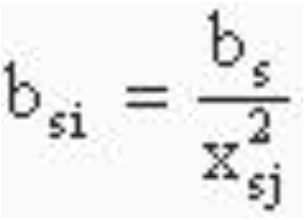 (3)
(3)
при всех s и i , i = s+1-j
Складывая по I обе части равенства (3) и положив
![]() (4)
(4)
получаем самое основное диофантовое уравнение, непараметрическое уравнение(-кроме параметра-аргумента размерности s ) в обратных квадратах:
![]() ,
(5)
,
(5)
которое, в добавок, является дискретной формой характеризационного требования нормировки функции распределения
![]() (6)
(6)
Отметим,
что в последних выражениях нумерация
по i
и по j
- взаимно обратная. Это определилось
нумерацией хsi
в решениях условно именуемыми нами
решениями В. Серпинского: при росте i
хsi
растет, а bsi
уменьшалось бы при выборе bsi
![]() 1/ xsi2
, а это было сочтено нецелесо-образным,
поскольку считалось естественным ходом
1/ xsi2
, а это было сочтено нецелесо-образным,
поскольку считалось естественным ходом
нумерации вариант, когда росту i отвечает рост bsi .
Наконец, дадим необходимые определения из теории сложности алгоритмов [30, Л-1] и согласно [7, Л-1] прида-дим им биологический смысл.
5. За относительную сложность кn (y,X) объекта y по отношению к заданному объекту х принята минимальная длина - целое число - l(p) программы p получения у из х, т.Е. К l(p).
6.
Если конечное подмножество M![]() N
(N-
множество с очень большим числом
элементом) допускает определение при
помощи программы длины пренебрежимо
малой по сравнению с loq2N,
то почти все элементы - объекты п.5 -
имеют сложность КN(y,x)
, близкую к loq2N.
Именно это утверждение формально роднит
понятие сложности алгоритма с традиционным
определением информации.
N
(N-
множество с очень большим числом
элементом) допускает определение при
помощи программы длины пренебрежимо
малой по сравнению с loq2N,
то почти все элементы - объекты п.5 -
имеют сложность КN(y,x)
, близкую к loq2N.
Именно это утверждение формально роднит
понятие сложности алгоритма с традиционным
определением информации.
7. Здесь по [7, Л-1] N отождествляется с числом всех элементарных актов хsi . а х и у - с описанием состояния растущей популяции: х - с начальным состояниями при s=1, а у -с уsj. , рассматриваемое как конечное.
8. Программу А(р, х) и соотношение А(р, х)=у, реализую-щее асимптотически оптимальный метод программиро-вания, отождествляем с процессом дифференцировки как в целом, так и на отдельных стадиях s,j, рассматриваемых как конечное. А(р, х), х, у, р - все величины снабжаются индексами s,j. 9. Из пп. 1-8 следует:
![]() (7)
(7)
Т.е. с помощью этой цепи соответствий показана взаимная непротиворечивость выше сообщенной кон-цепции «экономного» языка и понятия сложности алгоритма.