

Оттеснение рецессивов на окраину ареала
Аналогичное преобладание рецессивных аллелей на окраинах ойкумены можно проследить не только для пигментации, но и для многих других признаков, наследственно детерминированных малым числом генов. Прежде всего следует обратиться к такой полно изученной системе ге- нов, как группы крови АВО. Предложенная еще в середине 20-х годов Ф. Бернштейном гипотеза наследования этих групп пользуется всеобщим признанием.
Определение наследственных факторов крови системы АВО вошло в практику антропогенетических исследований с 1919 г. и насчитывает, таким образом, более полувека, за которые были изучены тысячи попу- ляций во всех, даже самых заброшенных, уголках земного шара. Соб- ранные данные позволили создать панойкумепные карты географического распределения аллелей, контролирующих систему АВО 60. На этих кар- тах отчетливо видны очаги концентрации п разрежения соответствующих аллелей, и они представляют неоценимый источник информации для по- нимания геногеографической природы тех закономерностей, которые управляют географическим распределением этих аллелей.
Рассмотрение много раз публиковавшейся карты рецессивного аллеля г (контролирующего наследование группы 0) обнаруживает совершенно отчетливую картину. Наименьшая концентрация этого аллеля падает на центральные районы Евразии. Предлагалось много гипотез о центрах возникновения аллелей группы АВО. Некоторые из этих гипотез имели исходным положением представление о самостоятельности центров
возникновения аллеля; авторы таких гипотез выделяли три первичные расы по группам крови: европеоидов и пегроидов с преобладанием груп- пы А, азиатских монголоидов с преобладанием группы В и американои- дов с преобладанием группы О. Ни одна из этих гипотез не могла, од- нако, обойтись без дополнительных предположений при объяснении концентрации соответствующих аллелей у многих народов земли. Про- тиворечит такая постановка вопроса и представлению о единстве наслед- ственного фактора, определяющего наследственную передачу системы А ВО. Между тем стоит лишь принять гипотезу возникновения всех аллелей этого гена в центральных районах Старого Света, дополнив ее затем геногеографической концепцией Н. II. Вавилова,— и надобность в каких-лпбо дополнительных предположениях отпадает. Наименьшая концентрация аллеля г в центре Евразии становится понятной, так как это очаг генообразования, в котором доминантные аллели преобладают. По мере удаления от этого очага во всех направлениях концентрация аллеля г постепенно возрастает. В Европе она велика на самом севере Скандинавии, в Ирландии, в прибрежных районах Западной Европы; она заметно подскакивает в Африке, особенно в центральных районах, где в условиях тропического леса предпосылки для изоляции и проявления ре- цессивного аллеля особенно велики.
Так же резко концентрация увеличивается в Северной Азии и в Ав- стралии. Причина этого увеличения ясна: по мере удаления от очага генообразования выщепление рецессивного аллеля происходит все с боль- шей интенсивностью. Особенно резко это заметно на Американском мате- рике, где его концентрация становится подавляющей. В большинстве популяций Центральной и Южной Америки, в наибольшей удаленности от очага генообразования. рецессивный аллель почти полностью вытеснил доминантные и концентрация его близка или даже составляет 100%. Таким образом, он занял то положение, какое не запимают в центре ойкумены даже доминантные аллели. Картина последовательного на- растания концентрации этого аллеля от центра к периферии настолько логична и правильна, настолько не содержит исключений, что трудно от- делаться от мысли: в географическом распространении аллелей системы АВО явление оттеснения рецессивов из центра генообразования играло решающую роль.
Система АВО — не единственная, к объяснению географии которой мсжно с успехом привлечь закон Н. И. Вавилова. Не менее демонстратив- но в этом отношении географическое распределение аллелей, детермини- рующих передачу по наследству групп крови Р. Ген образует два алле- ля — Р и р, из которых Р является доминантным. Концентрация этих аллелей изучена в меньшем числе популяций по сравнению с концентра- цией аллелей системы АВО, и потому карты их географического рас- пределения менее подробны. До сих пор публиковались лишь карты гео- графии аллеля Р6', но при автоматической зависимости, которая суще- ствует между концентрациями Р и р, карта аллеля р будет перевернутой копией уже опубликованных карт. Сейчас количество данных значитель- но увеличилось (рис. 4). Центр концентрации доминантного аллеля Р — Африканский материк. В Европе, Передней и Южной Азии удельный вес доминантного и рецессивного аллелей приблизительно одинаков. Но концентрация последнего резко возрастает в отдельных группах коренных австралийцев, по всей Новой Гвинее, у сибирских монголоидов, по всему северу Северной Америки. Поскольку рецессивный аллель нигде не ус- пел вытеснить доминантный полностью, как это было в системе АВО,
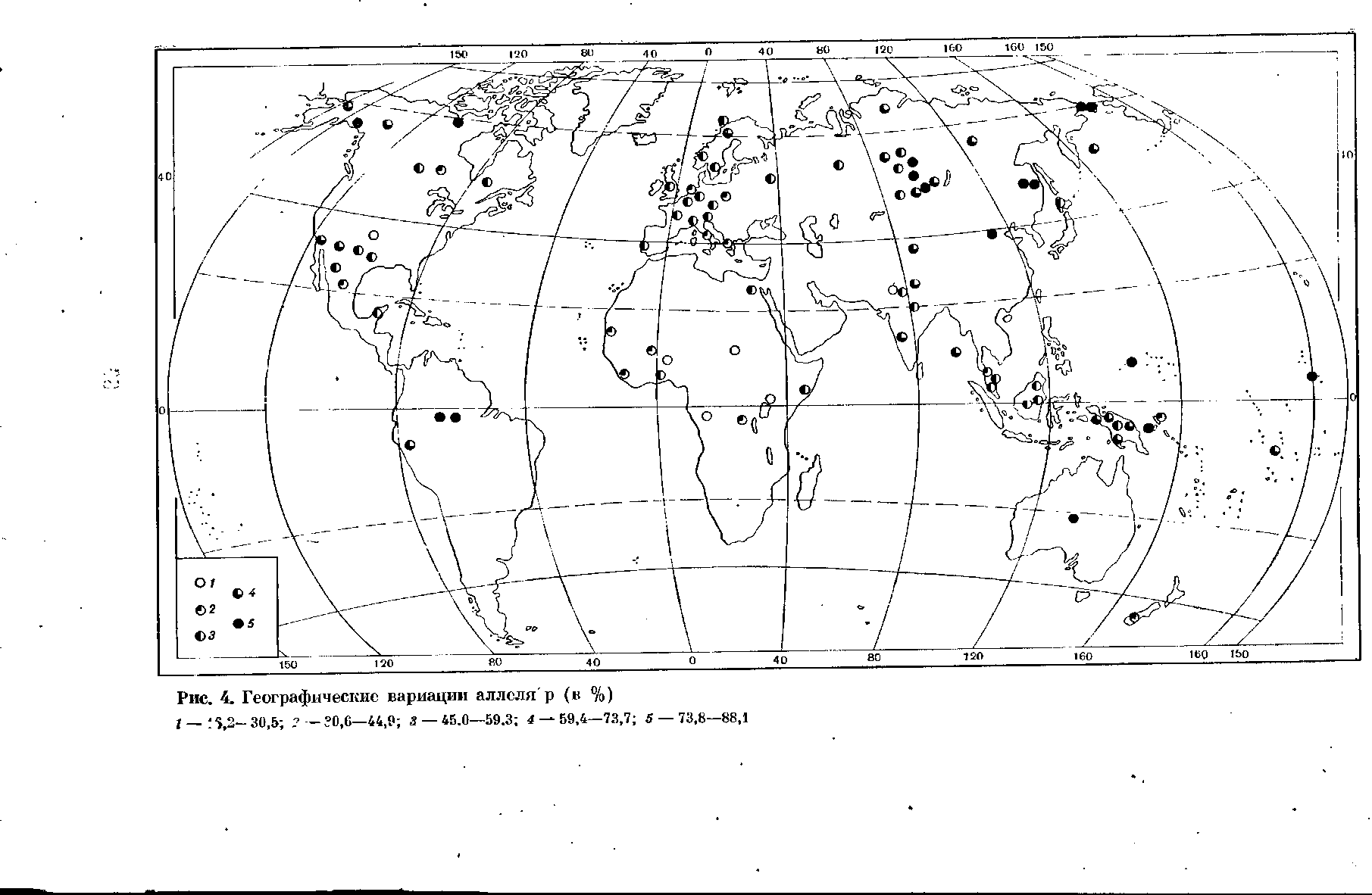
в центральных и южных районах Северной Америки, в Центральной Америке, у некоторых индейских групп Южной Америки концентрация рецессивного аллеля опять понижается, а концентрация доминантного по- вышается до уровня европейских величин.
По-видимому, в географии отдельных аллелей наряду с геногеографи- ческими закономерностями известную роль играет хронологический мо- мент, эпоха их появления, т. е. удаленность соответствующей мутации от современности, и время, необходимое на протекание тех или иных геногеографических процессов. Не исключено, что степень изоляции, в которой находились по отношению друг к другу индейские племена США и Центральной Америки, меньше, чем в Канаде, что при сохране- нии в достаточном проценте случаев доминантного аллеля Р ослабило темп выщепления рецессивного аллеля и повело к повторному увеличе- нию концентрации Р. К сожалению, мы пока лишены возмояшости су- дить сколько-нибудь точно о времени возникновения отдельных групп крови и вообще тех или иных вариаций физиологических признаков. Воз- вращаясь к конкретной географии группы крови Р, можно зато сказать с полной определенностью, что в условиях резкой, почти полной изоляции, в каких до недавнего времени находились таежные народы Сибири и до сих пор находится население Новой Гвинеи, концентрация рецессивного аллеля р резко подскакивает. Налицо, следовательно, та же закономер- ность оттеснения рецессива из центра генообразования и концентрация его на периферии.
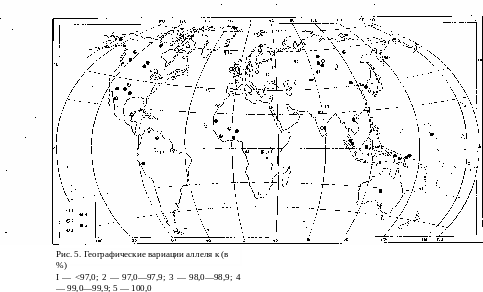
Передача по наследству системы группы крови Келл детерминирует- ся, как и Р, двумя аллелями, из которых К—доминантный и к — рецессивный. Концентрация доминантного аллеля даже в тех популя- циях, которые отличаются его максимальной частотой, очень певелика и составляет 4—5% (только в одном случае — 12%). В соответствии с этим велика, наоборот, вероятность случайной потери доминантного аллеля и замены его рецессивным. Эта вероятность во много раз больше, чем в предыдущих случаях, когда доминантные гены в центрах наибольшей своей концентрации были характерны для половины или большей части популяции. Все же и в этом случае географического распределения ре- цессивного аллеля обнаруживаются вполне отчетливые правильности62 (рис. 5). Наименьшая концентрация его падает на население Европы, где находится центр наибольшего уровня доминантного аллеля, а во всех направлениях от этого центра нарастает удельный вес рецессивного аллеля, в подавляющем большинстве популяций составляющего 100% генофонда популяции. Два исключения из этого правила—бушмены и индейцы Бразилии — не меняют общей картины, так как геногеографи- ческие закономерности, как уже указывалось, представляют собой вы- ражение вероятностных статистических процессов, при которых неизбеж- ны отдельные случайные отклонения от основной тенденции.
География группы крови Лютеран изучена также не очень полно, как и только что разобранной группы крови Келл, что связано с их сравнительно недавним открытием. Все же общее представление о гео- графическом распределении аллелей гена, определяющего наследствен- ную передачу группы крови Лютеран, можно составить па основании уже опубликованных данных (рис. 6). Этих аллелей два — Lu° и Lub , из ко- торых первый доминантен. Его максимальная концентрация не превыша- ет 8—9%, и поэтому по отношению к нему остается справедливым все то, что сказано выше о возможности случайного исчезновения аллеля
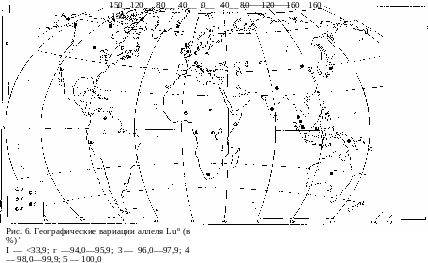
К. Центр концентрации аллеля Lu° — Европа и Африка, за исключени- ем южных районов. Население северных и центральных районов Азиат- ского материка, к сожалению, пока очень плохо изучено на предмет концентрации аллелей Lua и Lub. Но в Южной Азии и в Австралии, на юге Африки, в Северной и Южной Америке удельный вес рецессивного аллеля резко увеличивается, доходя во многих популяциях до полного, стопроцентного вытеснения им доминантного аллеля. Это опять случай значительного преобладания концентрации рецессивного аллеля на пе- риферии по сравнению с возможным центром гепообразовапия, где по- вышено, наоборот, относительное содержание в генофонде популяций до- минантной формы.
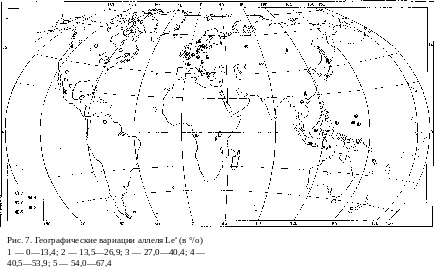
Система группы крови Льюис изучена географически еще более не- полно, чем две предыдущие. Карта географического распределения ал- лелей также напоминает предыдущие, но как бы в зеркальном отобра- жении (рис. 7): рецессивный аллель Lea концентрируется в Европе, тогда как по всей остальной ойкумене преобладает доминантный аллель, в Австралии и Америке достигая в своей концентрации 100%. На пер- вый взгляд здесь нарушается установленная Н. И. Вавиловым законо- мерность, но такое впечатление может быть и обманчивым, если учесть, что главным при рассмотрении правильностей в географическом распре- делении генов является не оценка центральных и периферических рай- онов ойкумены вообще, а выделение периферических районов по отноше- нию к центрам генообразования. Можно сделать предположение, что в последнем случае центром была территория, например, Южной и Юго- Восточной Азии. Оттуда исходная концентрация (100% или почти та- кая же концентрация доминантного аллеля) была перепесена в Амери- ку и Австралию. При стопроцентном преобладании домипаптного аллеля не создается, естественно, никаких предпосылок для выщепления рецес- снва в условиях изоляции просто в силу его отсутствия. Там же, где случайно он оказался представленным хотя бы в малом проценте слу- чаев в исходной группе, изоляция, даже относительная, сразу же при- водит к увеличению его удельного веса. Пример тому — даяки Борнео, яванцы и маори. Исходные группы, заселившие Европу, очевидно, также имели какой-то, пусть очень небольшой, процент рецессивного аллеля. Если рассматривать Южную и Юго-Восточную Азию как центр доми- нантных мутаций, то Европа в пределах Старого Света, конечно, пред- ставляет собой по отношению к этому центру периферию. Этим, по-ви- димому, и можно объяснить значительное и повсеместное в Европе повышение частоты рецессивного аллеля. О распределении его в Афри- ке в целом судить трудно из-за недостатка данных, но негры Западной Африки сближаются с европейскими народами.
Аналогичное рассуждение представляется вполне правомерным и для объяснения географического распределения аллелей группы крови Даффи (рис. 8). Если центром мутирования этого гена и, следователь- но, центром концентрации его доминантного аллеля считать Южную п Юго-Восточную Азию, то легко представить себе, что исходная кон- центрация была перенесена в Америку и Австралию, где и сохранилась до настоящего времени. Оттеснение рецессивного аллеля на периферию в Старом Свете вызвало значительное снижение частоты доминантного п увеличение концентрации рецессивного аллелей в Европе и особенно в Африке. В Африке оно еще более заметно, чем в Европе. Единствен- ным исключением из этого правила являются саамы на Крайнем Севере Европы, но следы их заведомо восточного происхождения проявляются
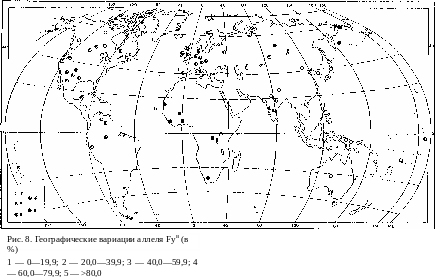
и в других признаках как морфологического, так и физиологического по- рядка. Не исключено, что некоторое понижение частоты рецессивного аллеля в Европе по сравпепию с Африкой также обязано своим проис- хождением более тесным контактам с восточными монголоидными попу- ляциями.
Данные о географическом распространении группы крови Кидд нельзя использовать из-за их неполноты и фрагментарности. География гена, определяющего наследственную передачу группы крови Диего, его преимущественная концентрация в доминантной форме в Централь- ной и Южной Америке свидетельствуют о его сравнительно позднем возникновении в пределах Америки, уже после переселения значитель- ных масс населения на Американский материк. В добавление ко всему уже сказанному о географическом распространении групповых факторов крови по земной поверхности и о подчинении его геногеографической закономерности, открытой Н. II. Вавиловым, можно привести еще один косвенный аргумент, отражающийся на географии системы MN и бел- ков сыворотки — гаптоглобинов. И группы крови системы MN, и гапто- глобпны имеют сходный тип наследования, будучи детерминированы двумя ко доминантными аллелями, т. е. аллелями, которые пе подавляют друг друга. В отношении этих систем нельзя поэтому говорить о доми- нантных или рецессивных аллелях. Очевидно, если предполагать влия- ние периферического положения этнических групп и изоляции на эти системы, его следует ожидать в увеличении доли одной из гомозигот пли обеих вместе. На географической карте аллелей гаптоглобина вид- но значительное увеличение процентной доли одного из аллелей в Се- верной Азии, в северных районах Северной Америки, на Новой Гви- нее 63. Наоборот, концентрация другого аллеля подскакивает на побе- режье Западной Африки, в центральных районах Африканского материка, в Центральной Америке. Всё это районы, изолированные либо геогра- фически,, либо, до педавнего времени, социально.
То же и с системой MN. Аллель m концентрируется в составе корен- ного населения обеих Америк, достигая максимума у индейцев юга США, Центральной Америки, северных государств Южной Америки. Макси- мум концентрации аллеля п падает на коренное население Австралии, затем идет население Африки. Все остальные районы земного шара с развитой городской цивилизацией, с гораздо более интенсивными в целом связями между этническими группами занимают промежуточное положение между этими двумя полюсами.
Итак, закон Н. И. Вавилова применим не только к объяснению гео- графического распределения пигментации у человека, но и к географии групп крови, бурно развивающейся в последние 20—30 лет и давшей большое количество сведений, никогда, насколько мне известно, не рас- сматривавшихся в рамках этого закона. С помощью закона Н. И. Ва- вилова, как мы убедились, легко объясняются многие факты геногеогра- фии человека, и, более того, в этих фактах он получает новое, очень убедительное подтверждение. В высшей степенп перспективной пред- ставляется дальнейшая разработка этого закона на антропологическом материале.