

15. Изменение экосистем под влиянием антропогенного загрязнения биосферы. Основные принципы и законодательная база природоохранной деятельности
Загрязнение биосферы - это любое внесение в ту или иную экологическую систему не свойственных ей живых или неживых компонентов, физических или структурных изменений, прерывающих или нарушающих процессы круговорота и обмена веществ, потоки энергии со снижением продуктивности или разрушением данной экосистемы.
Различают природные загрязнения, вызванные природными, нередко катастрофическими, причинами, например извержение вулкана, и антропогенные, возникающие в результате деятельности человека.
Антропогенное загрязнение – один из движущих факторов техногенеза. Техногенез — процесс изменения экосистем под воздействием производственной деятельности человека. Он проявляется в преобразовании экосистем, вызываемом совокупностью геохимических процессов, которые связаны с технической и технологической деятельностью людей.
Антропогенные загрязнители делятся на материальные (механические, химические, биологические) и физические, или энергетические (тепловая энергия, электрические и электромагнитные поля, шум, вибрация и т. д.).
Можно выделить следующие виды влияния антропогенного загрязнения на экосистемы:
1. Увлечение концентрации веществ или мощности энергетических полей, вызывающих непосредственно или косвенное изменение химических и физических свойств среды. Например, за счет газов антропогенного происхождения образуются кислотные осадки (в результате выбросов оксидов азота и серы), смог (ядовитый туман, содержащий широкой спектр токсических и канцерогенных газов) и происходит разрушение озонового экрана (разрушение озона хлорфторуглеродами). Последнее ведет к изменению спектральных свойств солнечной радиации, поступающей к поверхности Земли. Шумовое и вибрационное загрязнения ведут к миграции животных. Радиационное загрязнение усиливает мутагенез.
2. Выброс веществ, ведущих к возникновению и усилению других видов загрязнения. Быстрыми темпами растет в атмосфере содержание углекислого газа и метана. Эти газы обусловливают «парниковый эффект». Они пропускают солнечный свет, но частично задерживают тепловое излучение, испускаемое поверхностью Земли. Таким образом, данные газы ведут к тепловому загрязнению биосферы. Другим примером является избыточное поступление биогенов в водоемы, что стимулирует несбалансированный рост фитопланктона. Данный процесс называют эвтрофизацией. То есть химическое загрязнение ведет к биологическому загрязнению. К одному из видов загрязнения природных вод относится и тепловое загрязнение. Промышленные предприятия, электростанции нередко сбрасывают в водоемы (водохранилища) подогретую воду, приводящую к повышению в них температуры. В водоемах с повышением температуры уменьшается содержание кислорода, увеличивается токсичность загрязняющих воду примесей, нарушается биологическое равновесие, происходит смена видового состава организмов, например, водорослей. То есть тепловое загрязнение приводит к биологическому и химическому загрязнениям.
Изменение физико-химических свойств биотопа экосистемы закономерно ведет к изменению структуры ее биоценоза. При этом с видами организмов возможны следующие процессы:
1. Миграция. Часть популяций может мигрировать, найти местообитания с подходящими условиями и продолжить там свое существование.
2. Адаптация. В генофонде могут присутствовать аллели, которые позволят отдельным особям выжить в новых условиях и оставить потомство. Через несколько поколений под действием естественного отбора возникает популяция, хорошо приспособленная к изменившимся условиям существования.
3. Вымирание. Если ни одна особь популяции не может мигрировать, опасаясь воздействия неблагоприятных факторов, а те уходят за пределы устойчивости всех индивидов, то популяция вымрет, а ее генофонд исчезает.
С начала XVII по конец XX в. с лица Земли исчезло 68 видов млекопитающих, 130 видов птиц, 28 видов рептилий, 6 видов рыбы и 6 видов амфибий.
По данным Международного союза охраны природы (МСОП), в среднем на нашей планете ежегодно исчезает по одному виду или подвиду позвоночных животных. Помимо полного и повсеместного вымирания видов, широкое распространение приобрело явление частичного вымирания, т. е. исчезновения в отдельных странах и регионах.
Если одни виды вымирают, а выжившие особи других размножаются, адаптируются и изменяются под действием естественного отбора, можно говорить о поступательном изменении экосистемы, которое, в конечном счете, приводит к смене одного биоценоза другим с иным набором господствующих видов, а следовательно и иной трофической структурой. Данные смены одного биоценоза другим называют экзогенетическими.
Антропогенное загрязнение приводит к постепенному упрощению структуры биоценоза, обеднению их состава, снижению продуктивности. Подобные смены экосистем называют дигрессиями. Примерами дигрессий в следствии антропогенного загрязнения является превращения озера в болото в результате эвтрофизации.
Закон эволюционно-экологической необратимости гласит: экосистема, потерявшая часть своих элементов или сменившаяся другой в результате дисбаланса экологических компонентов, не может вернуться к первоначальному своему состоянию в ходе сукцессии, если в ходе изменений произошли перемены в экологических элементах (сохранившихся или временно утерянных). При снижении видового разнообразия за критический уровень климакс идентичный прошлому, не может быть достигнут.
Принципы природоохранной деятельности установлены в 3 статье Федерального Закона «Об охране окружающей природной среды» (от 10.01.2002):
Хозяйственная и иная деятельность, оказывающая воздействие на окружающую среду, должна осуществляться на основе следующих принципов:
обеспечение благоприятных условий жизнедеятельности человека;
научно обоснованное сочетание экологических, экономических и социальных интересов человека, общества и государства в целях обеспечения устойчивого развития и благоприятной окружающей среды;
охрана, воспроизводство и рациональное использование природных ресурсов как необходимые условия обеспечения благоприятной окружающей среды и экологической безопасности;
платность природопользования и возмещение вреда окружающей среде;
независимость контроля в области охраны окружающей среды;
презумпция экологической опасности планируемой хозяйственной и иной деятельности;
обязательность оценки воздействия на окружающую среду при принятии решений об осуществлении хозяйственной и иной деятельности;
приоритет сохранения естественных экологических систем, природных ландшафтов и природных комплексов;
обеспечение снижения негативного воздействия хозяйственной и иной деятельности на окружающую среду в соответствии с нормативами в области охраны окружающей среды, которого можно достигнуть на основе использования наилучших существующих технологий с учетом экономических и социальных факторов;
сохранение биологического разнообразия;
запрещение хозяйственной и иной деятельности, последствия воздействия которой непредсказуемы для окружающей среды, а также реализации проектов, которые могут привести к деградации естественных экологических систем, изменению и (или) уничтожению генетического фонда растений, животных и других организмов, истощению природных ресурсов и иным негативным изменениям окружающей среды;
соблюдение права каждого на получение достоверной информации о состоянии окружающей среды, а также участие граждан в принятии решений, касающихся их прав на благоприятную окружающую среду, в соответствии с законодательством;
ответственность за нарушение законодательства в области охраны окружающей среды;
организация и развитие системы экологического образования, воспитание и формирование экологической культуры;
международное сотрудничество Российской Федерации в области охраны окружающей среды.
Законодательная база природоохранной деятельности в России представлена следующими основными законами:
КОНСТИТУЦИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ.
ОБ ОХРАНЕ ОКРУЖАЮЩЕЙ СРЕДЫ. Федеральный Закон от 10.02.02 №7-ФЗ.
ВОДНЫЙ КОДЕКС РФ от 16.11.1995 N 167-ФЗ (С ИЗМЕНЕНИЯМИ НА 30 ДЕКАБРЯ 2001 ГОДА)
О НЕДРАХ Федеральный Закон от 21.02.92. N 2395-1
О ЗАЩИТЕ ТЕРРИТОРИИ И НАСЕЛЕНИЯ ОТ ЧС ПРИРОДНОГО И ТЕХНОГЕННОГО ХАРАКТЕРА Федеральный Закон от 21.12.94. N 68-ФЗ
О ПРИРОДНЫХ ЛЕЧЕБНЫХ РЕСУРСАХ, ЛЕЧЕБНО-ОЗДОРОВИТЕЛЬНЫХ МЕСТНОСТЯХ И КУРОРТАХ. Федеральный Закон
О ЖИВОТНОМ МИРЕ. Федеральный Закон
О МЕЛИОРАЦИИ ЗЕМЕЛЬ. Федеральный Закон
ОБ ОСОБО ОХРАНЯЕМЫХ ПРИРОДНЫХ ТЕРРИТОРИЯХ. Федеральный Закон от 14.03.95 N 33-ФЗ.
О РАДИАЦИОННОЙ БЕЗОПАСНОСТИ НАСЕЛЕНИЯ. Федеральный Закон
О КОНТИНЕНТАЛЬНОМ ШЕЛЬФЕ РФ. Федеральный Закон от 23 октября 1995 года
ОБ ЭКОЛОГИЧЕСКОЙ ЭКСПЕРТИЗЕ. Федеральный Закон от 3.04.96 N 174-ФЗ. 23 ноября 1995 года
ЛЕСНОЙ КОДЕКС РФ. Федеральный Закон
О БЕЗОПАСНОСТИ ГИДРОТЕХНИЧЕСКИХ СООРУЖЕНИЙ. Федеральный Закон от 21.07.97 №117-ФЗ
О ПЛАТЕ ЗА ПОЛЬЗОВАНИЕ ВОДНЫМИ ОБЪЕКТАМИ. Федеральный Закон от 06.05.98 №71-ФЗ
ОБ ОТХОДАХ ПРОИЗВОДСТВА И ПОТРЕБЛЕНИЯ. Федеральный Закон от 24.6.1998 N 89-ФЗ
О САНИТАРНО - ЭПИДЕМИОЛОГИЧЕСКОМ БЛАГОПОЛУЧИИ НАСЕЛЕНИЯ. Федеральный Закон от 30.03.1999 N 52-ФЗ
ОБ ОХРАНЕ ОЗЕРА БАЙКАЛ. Федеральный Закон
ОБ ОХРАНЕ АТМОСФЕРНОГО ВОЗДУХА. Федеральный Закон от 4.05.1999 N 96-ФЗ
О ЗЕМЛЕУСТРОЙСТВЕ. Федеральный Закон от 18.06.2001 N 78-ФЗ
16. Клеточная теория – одно из общепризнанных обобщений, утверждающих единство принципа строения и развития всех живых организмов, базирующегося на клетке как общей единице строения и функционирования.
Основные положения клеточной теории:
1) Клетка – элементарна единица живого. Клетка – это ограниченная активной мембраной, упорядоченная структурированная система биополимеров (белков, нуклеиновых кислот) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом. Клеточная структура является главной, но не единственной формой существования жизни. Неклеточными формами жизни можно считать вирусы. Правда, признаки живого (обмен веществ, способность к размножению и т.п.) они проявляют только внутри клеток, вне клеток вирус является сложным химическим веществом. По мнению большинства учёных, в своём происхождении вирусы связаны с клеткой, являются частью её генетического материала, "одичавшими" генами.
2) Клетка – едина система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование, состоящее из сопряженных функциональных единиц – органелл или органоидов.
3) Клетки сходны – гомологичны – по строению и по основным свойствам. Известно, что существует два типа клеток - прокариотические (клетки бактерий и архебактерий), не имеющие отграниченного мембранами ядра, и эукариотические (клетки растений, животных, грибов и протистов), имеющие ядро, окружённое двойной мембраной с ядерными порами. Между клетками прокариот и эукариот существует и множество иных различий. У большинства прокариот нет внутренних мембранных органоидов, а у большинства эукариот есть митохондрии и хлоропласты. В соответствии с теорией симбиогенеза, эти полуавтономные органоиды - потомки бактериальных клеток. Таким образом, эукариотическая клетка - система более высокого уровня организации, она не может считаться целиком гомологичной клетке бактерии (клетка бактерии гомологична одной митохондрии клетки человека). Гомология всех клеток, таким образом, свеодится к наличию у них замкнутой наружной мембраны из двойного слоя фосфолипидов (у архебактерий она имеет иной химический состав, чем у остальных групп организмов), рибосом и хромосом - наследственного материала в виде молекул ДНК, образующих комплекс с белками. Это, конечно, не отменяет общего происхождения всех клеток, которое подтверждается общностью их химического состава.
4) Клетки увеличиваются в числе путем деления исходной клетки после удвоения ее генетического материала (ДНК): клетка от клетки.
5) Многоклеточный организм представляет собой новую систему, сложный ансамбль из множества клеток, объединенных и интегрированных в системы тканей и органов, связанных друг с другом с помощью химических факторов, гуморальных и нервных (молекулярная регуляция). Кроме клеток, морфо-функциональными единицами тканей могут выступать надклеточные структуры – синцитии и симпласты – и постклеточные структуры: эритороциты большинства млекопитающих, тромбоциты.
6) Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией (работой) различных генов, что приводит к их морфологическому и функциональному разнообразию – к дифференцировке.
Морфо-функциональные особенности структурных компонентов эукариотической клетки:
Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Ядерная пора – это сложный белковый комплекс, имеющий окатэдрическую структуру, который активно регулирует ядерно-цитоплазматический транспорт.
Поверхностный комплекс животной клетки состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной. Это биологическая мембрана, толщиной около 10 нанометров. Обеспечивает в первую очередь разграничительную функцию по отношению к внешней для клетки среде. Кроме этого она выполняет транспортную функцию. На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции. Плазматическая мембрана животных клеток в основном состоит из фосфолипидов и липопротеидов со вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов. В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета— упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами» и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Эндоплазматический ретикулум
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубок и цистерн), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к гладкому (или агранулярному) ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПР не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Аппарат Гольджи
Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Лизосомы
Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной. В ней находятся литические ферменты, способные расщепить все биополимеры. Основная функция — автолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки.
Цитоскелет
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые, промежуточные филаменты и центироли. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3.
Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей.
Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек.
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий.
Внутренний просвет митохондрий, называемый матриксом отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки, так называемые кристы. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии.
Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что безусловно указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов. Самое большое число митохондриальных генов (97) из изученных организмов имеет простейшее Reclinomonas americana.
Пластиды – двумембранные органеллы, наружная мембрана которых гладка, а внутренняя образует замкнутые образования – тилакоиды. Наиболее развита система тилакоидтов у хлоропластов, где они образуют упорядоченные группы – граны. У лейкопластов, хромопластов и этиопластов тилакоидов меньше, и они располагаются беспорядочно. Пластиды способны нести фотосинтетические пигменты – хлорофиллы, которые входят встроены в супермолекулярные комплексы – фотосистемы, интегрированные в мембраны тилакоидов. Такие пластиды называются хлопластами. В них осуществляется фотосинтез. Кроме того, в их матриксе располагается активный ферментативный комплекс – цикл Кальвина. Хромпласты – это «старые» хлоропласты с разрушенным до каратиноидов хлорофиллом и деградировавшй мембранной структурой. Лейкопласты – специализированые пластиды на накопления вторичного крахмала (амилопласты), жиров (элайопласты) и белков (протеинопласты, или алейроновые зерна).
17. Обмен веществ — важнейшее свойство живых организмов. Совокупность реакций обмена веществ, протекающих в организме, называется метаболизмом. Метаболизм состоит из реакций ассимиляции (пластического обмена, анаболизма) и реакций диссимиляции (энергетического обмена, катаболизма). Ассимиляция — совокупность реакций биосинтеза, протекающих в клетке, диссимиляция — совокупность реакций распада и окисления высокомолекулярных веществ, идущих с выделением энергии. Эти группы реакций взаимосвязаны: реакции биосинтеза невозможны без энергии, которая выделяется в реакциях энергетического обмена, реакции диссимиляции не идут без ферментов, образующихся в реакциях пластического обмена.
По типу обмена веществ организмы подразделяются на две группы: автотрофы и гетеротрофы. Автотрофы — организмы, способные синтезировать органические вещества из неорганических и использующие для этого синтеза или солнечную энергию, или энергию, выделяющуюся при окислении неорганических веществ. Гетеротрофы — организмы, использующие для своей жизнедеятельности органические вещества, синтезированные другими организмами. В качестве источника углерода автотрофы используют неорганические вещества (СО2), а гетеротрофы — экзогенные органические. Источники энергии: у автотрофов — энергия солнечного света (фотоавтотрофы) или энергия, выделяющаяся при окислении неорганических соединений (хемоавтотрофы), у гетеротрофов — энергия окисления органических веществ (хемогетеротрофы).
Большинство живых организмов относится или к фотоавтотрофам (растения), или к хемогетеротрофам (грибы, животные). Если организмы, в зависимости от условий, ведут себя как авто- либо как гетеротрофы, то их называют миксотрофами (эвглена зеленая).
В процессе биосинтеза белка выделяют два основных этапа: транскрипция — синтез РНК на матрице ДНК (гена) — и трансляция — синтез полипептидной цепи.
Транскрипция – биосинтез РНК на матрице ДНК. В результате образуется м-РНК, кодирующая аминокислотные последовательности белков, а также т-РНК и др виды, выполняющие структурные, регуляторные и каталитические функции. В основе транскрипции лежит принцип комплементарности азотистых оснований цепей ДНК и РНК, а сам процесс осуществляется с участием соответствующих ферментов – РНК-полимераз и группой белков-регуляторов транскрипции. Детально изучены этапы механизма регуляции транскрипции прокариотических геномов, а эукариот мало. Транскрипции обычно подвергается одна из цепей ДНК, которая носит название матричной цепи (матрицы 3/-5/). Транскрипции подвергаются отдельные участки молекулы ДНК – транскриптоны. Они ограничены 2-мя последовательностями которые называются промотор (начало) и терминатор (остановка транскрипции). У бактерий транскриптоны называют оперонами (состоят обычно из нескольких цистронов (структурных генов), т. е. нуклеотидных последовательностей кодирующих структуру нескольких белков). Т.о на оперонах бактерии м-РНК является полицистроновой и может быть использована для синтеза нескольких белков в отличие от моноцистроновых м-РНК высших организмов. Белки-регуляторы способны связываться с определенными нуклеотидными последовательностями ДНК – энхансерами (усилителями).
Транскрипция у прокариот из 3-х этапов: 1. Инициация – происходит в зоне промотора, где находится старт-сигнал представляющий определенную последовательность Прибнова (на смысловой цепи) и на нее ориентируется холофермент РНК-полимеразы, происходит открытие промотора и образование открытого комплекса. 2. Элонгация цепи – после присоединения 8 рибонуклеотидов, присоединяется фактор элонгации и продолжается дальнейшее удлинение цепи. 3. Терминация – определяется особой нуклеотидной последовательностью в ДНК в зоне терминатора (ГЦ- богатый участок, представляющий собой палиндром (перевертыш), образуется шпилька, высвобождается синтезированная РНК, отсоединяется РНК-полимераза и восстанавливается двуспиральная структура ДНК.
Посттранскрипционный процессинг (созревание РНК). Молекулы м-РНК у прокариот не подвергаются процессингу (у некоторых транскрипция и трансляция сопряжены). Т-РНК содержат минорные основания, образующие петли. Р-РНК тоже минорные основания.
Транскрипция у эукариот – участвуют 3 РНК-полимеразы (1-в ядрышке (синтез р-РНК), 2 – нуклеоплазме (синтез м-РНК), 3 – нуклеоплазме (т-РНК). Те же этапы, но другие ферменты и последовательности оснований, чем у прокариот. М-РНК подвергается процессингу (модификация и сплайсинг (вырезание интронов) и соединение экзонов. Процессинг т- и р-РНК аналогичен прокариотам.
Трансляция – декодирование м-РНК в результате которого информация с языка последовательности оснований мРНК переводится на язык аминокислотной последовательности белка. У прокариот стартовым сигналом к началу синтеза белка служит расположенный на матричной РНК кодон АУГ, кодирующий метионин. Этапы трансляции: 1. Инициация – начинается с образования инициирующего комплекса на 30 – S субчастице (к 5/ концу м-РНК), состоящего из м-РНК, 30 – S субчастицы рибосомы и молекулы фарминметионин-т-РНК, которая связывается с кодоном АУГ. Далее присоединяется 50-S субчастица и образуется 70- S инициирующий комплекс. Участвуют еще факторы инициации. Используется энергия гидролиза ГТФ. 2. Элонгация – последовательное включение аминокислотных остатков в состав растущей полипептидной цепи. Каждый акт элонгации из трех этапов: узнавание кодона (т-рнк узнает), образование пептидной связи, транслокация. 3. Терминация – факторы терминации узнают терминирующие кодоны на аминоацильном сайте. Происходит гидролиз связи между концевым пептидом и т-РНК, свободный пептид диффундирует от рибосом. Высвобождается матричная РНК от рибосомы, которая распадается на 30S и 50S субчастицы.
Механизмы инициации трансляции у эукариот. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
У эукариот существуют два механизма нахождения рибосомой стартового AUG: кэп-зависимый (сканирующий) и кэп-независимый (внутренняя инициация).
При сканирующем механизме рибосома (точнее, её малая субъединица) садится на 5'-конец мРНК в области кэпа и двигается вдоль молекулы мРНК, «сканируя» один кодон за другим, пока не наткнётся на инициаторный AUG. Для привлечения рибосомы к 5'-концу мРНК требуется специальная структура, кэп — 7-метилгуанин, прикреплённый к 5'-концевому нуклеотиду мРНК.
При механизме внутренней инициации, называемом у эукариот также IRES-зависимым механизмом, рибосома садится на внутренний участок мРНК, называемый IRES (от англ. Internal Ribosomal Entry Site, участок внутренней посадки рибосомы) — участок мРНК, обладающий выраженной вторичной структурой, позволяющей ему направлять рибосомы на стартовый AUG. По IRES-зависимому механизму инициируется синтез лишь на небольшой части клеточных мРНК, а также на РНК некоторых вирусов.
Реакции матричного синтеза
Это особая категория химических реакций, происходящих в клетках живых организмов. Во время этих реакций происходит синтез полимерных молекул по плану, заложенному в структуре других полимерных молекул-матриц. На одной матрице может быть синтезировано неограниченное количество молекул-копий. К этой категории реакций относятся репликация, транскрипция, трансляция и обратная транскрипция (синтез ДНК на матрице РНК).
18. Изменчивость – способность организма приобретать или утрачивать определенные признаки и свойства. Делится на : генотипическую (мутации и комбинации), фенотипическую(или модификационная) и промежуточное положение занимает онтогенетическая.
Генотипическая связана с изменением генотипа, поэтому она наследственная. Комбинативная изменчивость – изменение фенотипа связанное с изменением генотипа на основе: 1. Свободного комбинирования генов (жел. гладкие дали жел. морщинистые); 2. На основе взаимодействия генов (эпистаз (когда один доминантный неаллельный ген подавляет действие другого неаллельного гена), криптомерия (когда рецессивная пара аллелей оказывает ингибирующее действие, еще называют рецессивный эпистаз), комплементарный тип взаимодействия (доминантные неаллельные гены при совместном действии приводят к новообразованию по сравнению с действием каждого гена в отдельности (окраска оперения у попугаев)); 3. На основе рекомбинации генов (серое тело нормальные крылья стали серые с зачаточными крыльями). Поэтому комбинативная изменчивость не является внезапной и прогнозируема.
Мутации – скачкообразное изменение фенотипа связанное с изменением генотипа. Свойства мутаций: 1. Дискретны (прерывисты); 2. Внезапны, скачкообразны; 3. Наследственны; 4. Не направлены (изменяться может любой локус гена как жизненноважный так и нет); 5. Повторяемость – одни и те же мутации могут возникать неоднократно; 6. Индивидуальны (индивидуальностью генотипа); 7. Редки (на 10000 экземпляров генов 1 мутация). Мутации по изменению генотипа делятся на генные, хромосомные, геномные и цитоплазматические.
Генные мутации – изменение пар нуклеотидов ДНК или РНК (трансцизии (замена пар нуклеотидов, которые не изменяют общей пуриново-пиримидиновой ориентации в пределах пары), трансверсии (изменяют), вставка лишней пары, выпадение пары нуклеотидов). Причина возникновения по концепции Борстелла это «ошибки 3R» - рекомбинации, репликации и репарации. «Генные болезни» - катаракта, гемофилия, дальтонизм, астигматизм, арахнодактилия. Адаптивная роль – увеличивают резерв мутационной изменчивости.
Хромосомные – изменение структуры целых генов (аберрации) или блоков генов (1. Нехватки: а) дефишенси или концевая б) делеция или срединная При нехватках нарушается конъюгация и кроссинговер («запиратели кроссинговера»); 2. Дупликации (добавление одного или блоков генов) а)терминальные б)срединные в)тандемные г)инвертированные (перевернутые) д)мультипликации (множественные повторы) Создают дополнительные участки генетического материала, функции которого могут меняться в ряду поколений, следовательно стабильность; 3. Инверсии (поворот блоков генов на 1800) 4. Транслокации (изменение положения генов или блоков генов) 5. Транспозиции – изменение локализации участков генетического материала с помощью мигрирующих генетических элементов – транспозонов (включают гены не имеющие отношения к перемещению) и инсерций (вставные последовательности, которые включают гены перемещения).
Геномные – изменение числа хромосом. Основаны на изменении генома. Делится на: 1. Полиплоидия – кратное гаплоидному набору : а) автополиплоидия - в пределах вида. (пример все махровые цветы (астры)) У растений происходит увеличение количественных показателей, но снижаются адаптивные качества (уменьшается устойчивость к заморозкам, заболеваниям); б)аллополиплоидия – умножение геномов разных видов (тритикале, слива, мул) и 2. Анеуплоидия – не кратное гаплоидному набору: а) 2n+1 трисомия (Синдром Дауна) б) 2n-1 моносомия (с. Шершевского-Тернера) в) 2n-2 нулесомия (искревление ствола, уродства)
Цитоплазматические – изменение плазмона (пример пестролистность).
Фенотипическая изменчивость – связана с изменением фенотипа под влиянием условий внешней среды и не затрагивает изменения генотипа. Свойства ф. изменчивости: 1. Не наследуется; 2. Носят групповой характер (под влиянием света все проростки оказываются зелеными); 3. Изменения соответствуют действию определенного фактора среды (окраска шерсти кроликов от температуры среды (светлая шерсть +300С, темная +20С), загар от УФ лучей); 4. Изменения обусловлены нормой реакции – предел варьирования признака, которая наследуется! Значение модификационной изменчивости: 1. Обеспечивает адаптации 2. Эволюционное (благодаря наследованию нормы реакции). Ф. и. делится на: 1. Адаптивные модификации – реакция клеток или организма на изменение условий среды, которые неблагоприятно действовали на организм в ходе эволюции (кол-во хлоропластов и хлорофилла у тене- и светолюбивых растений); 2. Морфозы – изменения при интенсивном действии факторов среды: а) фенокопии мутаций – напоминают фенотипическое проявление мутаций (заячья губа, волчья пасть) б) фенокопии нормы - напоминают фенотипическое изменение похожее на норму; 3. Длительные модификации – когда предварительное действие небольших доз ядов вызывает устойчивость к последующему действию больших доз; 4. Неспецифические адаптивные модификации (белки теплового шока); 5. Онтогенетические адаптации развивающегося организма.
Онтогенетическая изменчивость - наблюдается в процессе индивидуального развития (онтогенеза). О. и. представляет собой один из видов фенотипической изменчивости. Примеры о. и.: яйцевой зуб у крокодилов, средняя продолжительность жизни дрозофилы зависит от температуры выращивания мушек предыдущего поколения.
Спонтанный мутационный процесс – возникновение мутаций естественным путем в более или менее нормальных условиях. Причины «ошибки 3R» - рекомбинации,репликации, репарации. Обусловлено природными факторами: 1. Естественный фон радиации 2. Аутомутагены – метаболиты (перекись, спирт) 3. Резкие колебания температуры 4. Гены-мутаторы – их функция мутировать и повышают мутирование других генов 5. Инфекционный мутагенез – вирусы, бактерии, гельминты 6. Существующие в ДНК участки повторы 7. Инсерционные последовательности и транспозоны 8. Ошибки кодирования ДНК. Характеристики спонтанного м. п. : 1. Определенная частота возникновения (от общего числа генов у конкретной особи и от общего числа особей в популяции) 2. Находится под генетическим контролем (разные гены в одном генотипе могут мутировать с разной скоростью и частотой) 3. Мутабильность – относительная способность гена мутировать (гены мутаторы есть у всех).
Индуцированный м. п. – с участием экспериментатора с участием мутагенов (агентов окружающей среды, вызывающих мутации): 1. физические – ядерная энергия, УФ лучи, ионизирующее излучение, переменное магнитное поле; 2. химические – пестициды, нитраты, отходы, пищевые добавки; 3. Биологические – вирусы, бактерии, возраст, стресс.
Согласно одной из наиболее поздних классификаций, антимутагены делятся на две большие группы — внеклеточные (ингибиторы образования и поглощения мутагенов и ингибиторы превращения промутагенов в мутагены) и внутриклеточные (блокаторы поступления мутагенов в клетки, взаимодействующие с генетическими структурами за счет усиления активности детоксицирующих ферментов или прямого взаимодействия с мутагеном, антиоксиданты, т.е. ловушки свободных радикалов, и агенты, влияющие на репарацию генетических повреждений).
Методы тестирования мутагенов.
Была признана необходимость первоочередной проверки на мутагенность лекарств, пищевых добавок, пестицидов, гербицидов, инсектицидов, косметических средств, наиболее широко распространенных загрязнителей воды и воздуха, а также производственных вредностей. Второй методологический принцип заключается в выборочности тестирования (это означает, что вещество анализируется на мутагенность при наличии двух обязательных условий: распространенности в среде обитания человека и наличии структурного сходства с известными мутагенами или канцерогенами).
Отсутствие универсального теста, позволяющего одномоментно регистрировать индукцию изучаемым веществом (и его возможными метаболитами) различных категорий мутаций в половых и соматических клетках, служит основанием третьего принципа - комплексного использования специализированных тест-систем. Наконец, четвертый методологический принцип подразумевает ступенчатость тестирования веществ на мутагенную активность.
Этот принцип берет начало от одной из первых и наиболее известных схем, предложенной в 1973 г. Б, Бриджесом и предусматривавшей три последовательных этапа исследования.
1. На первом этапе мутагенные свойства вещества изучали простыми и быстро выполнимыми методами (с использованием микроорганизмов и дрозофилы в качестве тест-объектов) для определения его способности индуцировать генные мутации. Выявление такой способности предполагало запрет на применение данного вещества.
2. В случае особой медицинской или экономической значимости мутагена его тестировали на млекопитающих in vivo. Аналогичное исследование проводилось также для веществ, не продемонстрировавших мутагенных свойств в тестах первого этапа. Если исследуемый агент не проявлял мутагенных свойств, постулировалась безопасность применения его человеком. Вещества, проявившие мутагенность, либо запрещали для использования, либо, если они относились к категории особо значимых, или незаменимых, исследовали дополнительно. 3. На заключительном этапе проводили тестирование для установления количественных закономерностей мутагенного действия таких специфических веществ и оценку риска применения их человеком.
Данная схема послужила прототипом целого ряда методик комплексного тестирования на мутагенность (программу, предложенную в 1996 г. Дж. Эшби с соавторами). Исключительно важной особенностью этой программы является ее направленность не только на оценку мутагенности тестируемого вещества, но и на прогноз канцерогенности данного химического соединения и возможного механизма канцерогенеза.
В программе Дж. Эшби постулируется, что вещество не является канцерогеном, если оно не проявляет мутагенного и генотоксического действия in vivo. Те же вещества, которым названные эффекты свойственны, являются потенциальными генотоксическими канцерогенами.
Известно более 100 различных методов оценки генотоксичности. Однако реально практическое использование имеют не более 20 тест-систем.
Наиболее распространенный метод регистрации влияния ксенобиотиков (мутагенов и канцерогенов) на частоту генных мутаций предложен Брюсом Эймсом в 1975 г. Используемые в качестве тест-объекта His-мутанты Salmonella typhimurium не синтезируют гистидин и выживают на безгистидиновых средах только при возникновении обратной мутации к дикому типу His+. Ревертанты дикого типа образуют колонии на среде без гистидина, что и служит показателем возникновения генных мутаций. В последние годы тесг Эймса был значительно усовершенствован: автоматизирована процедура тестирования, повышена чувствительность к отдельным типам мутагенов.
Принципиально новый подход к оценке генотоксичности - использование трансгенных мышей с интегрированными в геном тестерными генами. По их изменению, можно оценивать индукцию генных мутаций. Используя трансгенные тест-объекты, можно изучать тканевую и органную специфичность мутагенного действия.
В большинстве случаев у высших организмов мутации отдельных генов не рассматриваются, поскольку они очень редки. Вместо этого оценивают частоту возникновения мутаций в хромосоме в целом.
Первый такой метод обнаружения и определения частоты мутаций у дрозофилы -СlВ— применил в (927 г. Герман Меллер. Он разработал систему, позволяющую отличать вновь возникшую мутацию от уже имеющихся в генотипе. В настоящее время метод С1В утратил свое практическое значение (между половыми хромосомами иногда может происходить двойной кроссинговер у самок). Вместо него используется предложенный в дальнейшем тем же автором метод Меллер-5. Метод Меллер-5, как и тест Эймса, широко применяется для контроля химических соединений, используемых при консервировании пищевых продуктов, изготовлении косметических препаратов и т.п.
Для оценки способности агентов индуцировать хромосомные мутации широко используются цитогенетические методы учета хромосомных аберраций. Недостаток этих методов состоит в том, что они достаточно субъективны (поскольку основаны на микроскопировании), требуют высокой квалификации исследователя и плохо поддаются автоматизации.
В качестве альтернативы был предложен метод учета микроядер (образуются из-за дефектов веретена деления) в эритроцитах костного мозга грызунов, который может быть автоматизирован и применен к любой пролиферируюшей ткани, вкчючая гонады.
Очевидно, что максимальное приближение исследователя к оценке генетического риска вследствие действия мутагенов внешней среды возможно только при использовании в качестве тест-систем клеток человека. В таких экспериментах обычно используются лимфоциты периферической крови и, как возможный вариант - клетки костного мозга, эпителия волосяных фолликулов, а также эмбриональные фибробласты и сперматозоиды (отсутствие единого стандарта в этой процедуре - различия в уровне доз, путей введения, сроков экспозиции). ВОЗ «Руководство по краткосрочным тестам для выявления мутагенных и канцерогенных химических веществ».
Тем не менее, пока в большинстве случаев оценка генетического риска основана на экстраполяции экспериментальных данных от одного тест-объекта на другой, от высоких доз на низкие и т.д., и, в конечном итоге, от модельных систем in vitro/in vivo — на человека.
Практически все исследователи считают проблему количественной экстраполяции чрезвычайно затрудненной. Причиной тому являются видовые, возрастные и индивидуальные особенности метаболизма. Метаболические особенности человека могут существенно повышать или понижать мутагенный эффект химических соединений. Следовательно, для правильной экстраполяции необходимо знать метаболизм конкретного мутагена и у животных, и у человека.
В зависимости от состава сравниваемых пар индуцированный мутагенез у человека может быть выражен сильнее или слабее. Например, показано, что человек более устойчив к мутагенному действию радиации, чем мышь. Не исключено, что таково же соотношение и в случае действия химических мутагенов. Существование количественных различий в результатах индуцированного радиационного мутагенеза связано с видовыми особенностями функционирования репарационных систем, которые способны восстанавливать первичные повреждении, имеющие потенциальный характер и не обязательно реализующиеся в мутации.
Ф. Собелсом сформулирован принцип экстраполяции, получивший название «правило параллелограмма» (допустимо применять строго при условии линейной зависимости эффекта от дозы).
19. Размножение (репродукция) - присущая всем живым существам функция воспроизведения себе подобных. Размножение направлено не на поддержание жизни отдельной особи, а на сохранение ее генов в потомстве и продолжение рода - тем самым на сохранение генофонда популяции, вида, семейства и т.д. Все разнообразие способов размножения можно разделить на два основных типа: бесполое и половое размножение.
Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Поддерживает существование вида, сохраняя его наследственные особенности. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение.
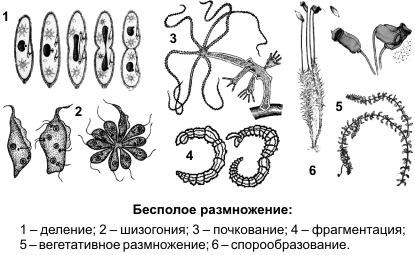
Деление — характерно для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить: а) простое бинарное деление (прокариоты), б) митотическое бинарное деление (простейшие, одноклеточные водоросли), в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов.
Почкование —новые особи образуются в виде выростов на теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
Фрагментация (4) —новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония —новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение —новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.).
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
Биологическая роль бесполого размножения
Поддержание наибольшей приспособленности в малоизменяющихся условиях окружающей среды. Оно усиливает значение стабилизирующего естественного отбора; обеспечивает быстрые темпы размножения; используется в практической селекции. Бесполое размножение встречается как у одно-, так и у многоклеточных организмов. У одноклеточных эукариот бесполое размножение представляет собой митотическое деление, у прокариот – деление нуклеоида, у многоклеточных форм – вегетативное размножение.
Половое размножение
Половое размножение осуществляется при участии двух родительских особей (мужской и женской), у которых в особых органах образуются специализированные клетки — гаметы. Процесс формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз. Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния мужской и женской гамет. Процесс слияния мужской и женской гамет называется оплодотворением. Обязательным следствием полового размножения является перекомбинация генетического материала у дочернего поколения.
В зависимости от особенностей строения гамет, можно выделить следующие формы полового размножения: изогамию, гетерогамию и овогамию.
Изогамия — гаметы являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия — женские и мужские гаметы являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия — женские гаметы неподвижные и более крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями. Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет. Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных организмов. При конъюгации происходит слияние содержимого отдельных гаплоидных клеток нитевидных талломов. По специально образующимся каналам содержимое одной клетки перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также делится мейозом.
Значение полового размножения. Оно обеспечивает значительное генетическое разнообразие и, следовательно, большую фенотипическую изменчивость потомства; организмы получают большие эволюционные возможности, возникает материал для естественного отбора.
Клеточный цикл (жизненный цикл) — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Значение митоза: 1. Осуществляется рост любого организма; 2. Регенерация тканей (репаративная и физиологическая); 3. Бесполое размножение.
Сущность митоза: 1. Всоздании точной копии генетического материала исходной клетки; 2. Равноценное распределение генетического материала по дочерним клеткам.
Митоз включает:
1. Аутосинтетическая интерфаза:
а) G1 период (накапливается энергия АТФ, синтезируются белки);
б) S период (синтезируется 2-я хроматида, репликация ДНК (у эукариот 3 способами: консервативным, полуконсервативным и дисперсным))
в) G2 период (снова накапливается АТФ, синтезируются специфические белки, необходимые для собственно митоза);
2. Собственно митоз:
а) Профаза (хромосомы подготавливаются к делению, спирализуются, конденсируются и становятся тельцами (чтобы перемещаться); ядрышко компактизируется на хромосомах и переносится со спутничной хромосомой в зону ядрышкового организатора; ядерная оболочка распадается на компоненты, оттесняется к мембране клетки; образуется веретено деления (центриоли расходятся к полюсам, а между ними микротрубочки, которые подходят к центромере); от каждого полюса к каждой хромосоме подходят две нити (тянущие));
б) Метафаза (хромосомы устанавливаются по экватору своими центромерами);
в) Анафаза (расщепляются центромеры (т.е. хромосомы расщепляются на хроматиды), затем хроматиды растягиваются к полюсам);
г) Телофаза (хромосомы деспирализуются, восстанавливается ядерная оболочка, формируется ядрышко, веретено деления частично разбирается, а остальное на построение цитоскелета, цитокинез (разделение клеточного тела)).
Мейоз – период созревания половых клеток, в результате которого половые клетки получают гаплоидный набор хромосом. Значение мейоза: повышает жизнеспособность организма, т.к. при оплодотворении восстанавливается диплоидный набор хромосом, что приводит к комбинативной изменчивости и возможен кроссинговер.
Мейоз представляет собой 2 деления следующих друг за другом:
Первое (редукционное) деление мейоза:
Интерфаза (как при митозе)
Профаза (тоже + кроссинговер)
Метафаза (тоже, но от каждого полюса идет нить к одной хромосоме)
Анафаза (хромосомы не расщепляются на хроматиды, а расходятся целыми, т. е. кол-во уменьшается вдвое);
Телофаза (хромосомы не деспирализуются, т.к. идет еще 2-ое деление, ядерная оболочка не строится, ядрышко при хромосоме, веретено не разрушается)
Второе эквационное деление – это типичный митоз, в интерфазу не происходит репликации ДНК, т.к. вторая хроматида уже есть.
Типы мейоза: 1. Зиготический (у водорослей) 2. Гаметный (у животных и человека при формировании половых клеток в яичниках и семенниках) 3. Промежуточный (у высших цветковых растений, в промежутке между формированием спор). Процесс мейоза универсальный.
Амитоз (термин предложен гистологом В. Флеммингом) - прямое деление клетки, при котором ядро находится в интерфазном состоянии. Спирализация хроматина не происходит, хромосомы не выявляются. Веретено деления не образуется. Амитоз приводит к появлению двух клеток, но очень часто в результате амитоза возникают двуядерные и многоядерные клетки. При амитозе делится только ядро, причем без образования веретена деления, поэтому наследственный материал распределяется случайным образом.
Амитотическое деление начинается с изменения формы и числа ядрышек. Крупные ядрышки делятся перетяжкой. Вслед за делением ядрышек происходит деление ядра. Ядро может делиться перетяжкой, образуя два ядра, или имеет место множественное разделение ядра, его фрагментация. Ядра могут быть неравной величины.
Амитоз встречается в отживающих, дегенерирующих клетках, неспособных дать новые жизнеспособные клетки.
В норме амитотическое деление ядер встречается в зародышевых оболочках животных, в фолликулярных клетках яичника.
Амитотически делящиеся клетки встречаются при различных патологических процессах (воспаление, злокачественный рост и др.).
20. Наследственность – способность организма передавать из поколения в поколение свои признаки и свойства. Материальной основой наследственности являются гены. Ген – участок любой генетической нуклеиновой кислоты (ДНК или РНК) несущий информацию о синтезе полипептидной цепи белковой молекулы или об элементарном свойстве организма или о синтезе функциональных РНК (т-РНК, р-РНК). Т. о. молекулярная основа наследственности заключается в особенностях строения НК: ДНК и РНК. В основе мономеры – нуклеотиды, которые состоят из 3-х составных частей: остаток фосфорной кислоты, сахар (дезоксирибоза и рибоза), пуриновые и пиримидиновые азотистые основания (А, Г, Т, Ц, У). Отличительная особенность – ДНК двуцепочечная, РНК одноцепочечная. Открыты НК Фридрихом Мишером. Согласно модели Крика – Уотсона, ДНК представляет двойную спираль. Двуспиральность определяет ее уникальность. Цепи ДНК – антипараллельны, шаг спирали составляет 3,4 нм, ширина спирали — 2 нм, длина спирали — несколько десятков тысяч нанометров. Специфичность ДНК определяется последовательностью азотистых оснований в ее цепочке. Уникальные свойства молекулы ДНК: 1. Способность к репликации (самовоспроизведению) 2. Видовая специфичность (у особей одного вида структурная организация ДНК одинакова) 3. Самокомплементарность (пуриновые и пиримидиновые основания).
Доказательством генетической роли НК явилось изучение генетики м/о. Изучение организации генома у бактерий (оболочка из муреина – полисахарид, может быть липидный слой от лизиса) и у фагов (белковый капсид) (генетическая информация как в виде ДНК (Т-четные), так и в виде РНК). Изучение не только хромосомных кольцевидных ДНК бактерий, но и внехромосомных – это плазмиды (могут быть автономными, а могут – в интегрированном состоянии, но с ДНК хромосомой (их виды: F- половой фактор (определяют место рекомбинации), R - резистентность (устойчивость к антибиотикам), Col- за синтез белков колицинов (токсичные белки вызывающие гибель других бактерий, но саму бактериальную клетку в которой они производятся они не убивают).
Способы передачи генетической информации на примере бактерий: 1. Конъюгация – обмен генетической информацией благодаря ЦП мостику (2 мутантных штамма бактерий после облучения: 1-ый не способен синтезировать витамин биотин и АК метионин, а 2-ой не синтезировал АК треонин и лейцин. Всреду без 4-х этих веществ поместили клетки каждого штамма и обнаружили колонии в клетках которых находились гены, отвечающие за синтез всех 4-х веществ). 2. Трансформация – процесс передачи свободной ДНК из одной клетки в другую (2 типа колоний пневмококк: 1 – S имеют капсулу защищающую от фагоцитоза и они вирулентные; 2 – R без капсул и не вызывают заболевания. Действием высоких температур на S разрушили капсулу и поместили к R, заразили этой смесью мышей и наблюдали их гибель, т. к. произошла мутация R в S и R стал способен синтезировать капсулу). 3. Трансдукция – передача генетической информации посредством бактериофага (в U-образную воронку в сонование поместили бакфильтр, в одно колено 22А штамм бактерий – не имеют гена триптофана, в другое 2А штамм (имеют ген триптофан) + бактериофаг. Через некоторое время в 2А штамме были клетки с геном триптофана).
Эти доказательства позволили раскрыть и расширить концепцию «1 ген – 1 полипептид».
Классические представления: 1. Т.Морган в работе «Теория гена» ген как единица функции, мутации (ген меняется целиком) и рекомбинации (между целыми генами), он не делим, т. е. гипотеза «1ген – 1 признак». При изучении аллелизма увидели несоответствие (изменение состояния гена, он более сложно устроен); 2. Серебровский изучал ступенчатый алелломорфизм (оказалось, что разные мутантные аллели приводят к разным изменениям фенотипа), т.е. изучали строение гена по фенотипу; 3. «1 ген – 1 фермент» расширилась когда Бидл и Татум исследовали биохимические мутации у нейроспоры и определили что аллельные мутации затрагивают те же функции биосинтеза белка; 4. Работы Бензера – мутация и рекомбинация может происходить как между генами, так и внутри них. Предложил мутон – единица мутации, рекон – рекомбинации, цистрон – единица функции (неизменный участок); 5. Работы Ингрема доказал, что серповидно-клеточная анемия наследуется как моногенный признак несмотря на то, что ген гемоглобина состоит из 2-х альфа и 2-х бета полипептидных цепей – поэтому возникла гипотеза «1 ген – 1 полипептидная цепь».
Современные представления о гене. Хромосома состоит из одной молекулы ДНК, содержащей множество генов. 95% генетической информации не транслируется, т. е. не кодирует последовательность АК полипептидной цепи молекулы белка, но транскрибируется, что позволило уточнить структуру гена. Основу транскрибируемых генов составляют структурные гены, которые имеют регуляторные участки и смысловую часть, которая у эукариот состоит из экзонов и интронов, а у прокариот из экзонов. Регуляторные участки делятся до смысловой (I) и после (II). В I части 2 группы: 1. Неспецифические последовательности – ТАТА –бокс, на расстоянии 30 нуклеотидных пар от смысловой, опознает РНК-полимеразу. Левее – промотор (сайт узнавания, связывания и инициации транскрипции). Перед промотором полиндром – является запирателем транскрипции в случае сдвига рамки считывания. 2. Специфические участки – энхансер, не сцеплен со смысловой и первой частью и находится на расстоянии десятков-сотен пар нуклеотидов; это первый специфический участок для РНК-полимеразы. Во II части находится терминальная последовательность завершающая транскрипцию.
У прокариот (Модель Жакоба и Моно) – ген –регулятор (несет информацию о белках-репрессорах), промотор (туда РНК-полимераза), оперон (оператор (к нему белок репрессор)+ группа структурных генов).
21. Физиология растительной клетки. Внутриклеточные механизмы регуляции. Регуляция обеспечивает гомеостаз организма, сохранение параметров внутренней среды и создание условий для развития растений. Т.к. первые ж. о. были одноклеточными, то первыми в ходе эволюции возникли внутриклеточные системы регуляции (мембранная, ферментативная, генетическая). Кл–основная структурная и функциональная ед-ца. Растительная клетка: оболочка и протопласт (гиалоплазма, органоиды, ядро); Органоиды: мембранные (одномембранные-ЭДС, вакуоль, АГ; двумембранные – пластиды, митохондрии) немембранные (рибосомы, микротрубочки). Мембранная регуляция. 1) обеспечивает увеличение рабочей поверхности; 2) деление клетки на компартменты (отсеки) в них специфические метаболические процессы. Отличит. особ-ти плазмолеммы раст кл-ки: 1) Обр-т плазмодесмы, каналы соед-ие соседние кл-ки. 2) испытывает давление со стороны цитоплазмы и кл стенки. Тонопласт-мембрана, огранич-ая вакуоль. Мембраны раст кл-ки имеют типичное липопротеидное строение. Наиб. приемлема жидкостно-мозаичная модель строения. Одним из важнейших свойств явл-ся динамичность-способность к движению белковых и липидных молекул, что позволяет адаптироваться к меняющимся условиям. Др. св-во- избирательная проницаемость-способность пропускать воду и определенные молекулы. (Доказательство-явление плазмолиза-потеря кл-ой воды при ее погружении в гипертонический раствор) Ф-ции мембран в кл-ке одновременно являются разными сторонами механизма регуляции внутриклет-ого обмена веществ. Особое значение имеет система мембранных хемо- фото- и механорецепторов, позволяющих кл-ке оценивать качественные и количественные изменения во внешней и внутренней среде и в соответствии с этим менять св-ва мембран. Мембранная регуляция осущ-тся путем сдвигов в мембранном транспорте, связыванию или освобождению ферментов и регуляторных белков, изменением активности мембранных ферментов. У эукариот с появлением ядерной обол-ки роль мембр регуляции возросла. Мембр. Регуляция контактная (ЭПР, АГ, где происх-т достройка и модификация секреторных белков, связывание мембр-ми освобожд-ся ферментов меняет их активность). Дистанционная (путем избирательной доставки ферментов, ионов, субстратов, удаление продуктов). Генетическая регуляция осущ-ся на уровне репликации и экспрессии генов (биосинтез белка): на уровне транскрибции и посттранскриционном уровне (процессинг, трансляция, созревание белка). Молекулярные механизмы осущ-ся за счет рН, активности ферментов, ионов. Не все гены в организме постоянно находятся в рабочем состоянии, существует сложная система регуляции. 1 гипотеза: 1961 Жакоб, Моно модель генетического контроля синтеза белков для прокариотической клетки. 2 гипотеза: 1997 Шарп и Робертс – гены эукариот это не непрерывные белоккодирующие нуклеотидн. последовательности, а мозаика из экзонов (кодирующие белки) и интронов (не кодирующие, несут регуляторную функцию). В геноме кроме структурных генов есть гены регуляторы, несущие информацию о белках регуляторах (их ф-ция включение/выключение гена). Белки регуляторы: 1 репрессоры (присоединяется к оператору не дает осущ-ть транскрибцию); 2 активаторы (способствуют транскипции). На способность репрессора соединяться с оператором влияют некоторые низкомолек-е в-ва (гормоны, аминокисл) –эффекторы. 2 вида: 1 корепрессор соед-ся с неактивной формой репрессора, переводит в активную, блокирует транскрипцию; 2индуктор соед-сь с репрессором мешают его способности взаимодействовать с оператором. Внутриклеточная регуляция осущ-ся так же за счет таких св-в цитоплазмы как вязкость, подвижность, эластичность.
Ферментативная регуляция. Ферменты-специфич белки, присутствующие во всех живых клетках, играющие роль биокатализаторов в р-циях обмена вещ-в. Св-ва: 1 катализир-т только термодинамически возможные р-ции 2 действуют в ничтожно малых кол-вах 3 не входят в состав конечных подуктов 4 Характерна специфичность: абсолютная- кат-т только 1 р-цию; относительная-синтезируют распад хим. связи в разных соединениях. Ферменты делят на простые (только из аминокислот) и сложные (белковая и небелковая части). Белковая-апофермент. Небелковая (в зависимости от прочности соединения с апоферм-ом) может быть: простетическая группа (прочно связана, атомы мет. Cu, Fe, Mg); кофермент (подвижно связан, витамины и их производные НАД, НАДФ, ФАД). Этапы работы: 1 образование фермент-субстратного комплекса (Фишер ключ-замок; Кошланд рука-перчатка) 2 Снижение энергии активации р-ции ( за счет деформации хим связей – смещение электронной пары) 3 выделение продуктов р-ции. Влияние условий: 1 концентрация ферм (чем больше, тем выше скорость) 2 концентрация субстрата (кривая Михаэлиса, сначала ск-ть р-ции увеличивается, затем не измен-ся, т к заняты все мол-лы фермента) 3 температура (термостабильные-теряют активность при 60о, термолабильные-выдерживают до 40о) 4 р-ция среды (узкий диапазон, б-во рН 7) 5 влияние атомов мет: +влияние а) входит в состав активного центра эф-т «металлические челюсти», б) за счет придания более opt конформации, эф-т «мет корсет»; - влияние: тяжелые мет проникая в мол-лу ферм нарушают структуру белка 6 фаза онтогенеза (сухие семена-мало ферм, замоченные-активация, появление новых).
Регуляция работы ферментов. Ферм м б конститутивными (постоянно присутствуют) и индуцированными (появл-ся иногда). 2 пути регулир-я активности ферм: 1) Мех-мы направл-е на поддержание клет-го гомеостаза 2) Мех-мы слежения за сигналами, поступ-ми в кл-ку. Они реализ-ся через след. Мех-мы: 1. Превращение неактивных предшеств-ов ферм (зимогенов) в активные ферменты; 2 Изостерический эф-т для низкомолек ферм, основан на конкурентном ингибир-ии (определенные в-ва конкурируют с субстратом за акт. Центр ферм) 3Аллостерический эф-т характ-н для крупных мол-л, имеющих дополнит сайт связывания с эффектором-аллостерический центр.
– эффекторы-некот фитогормоны, конечные прод р-ции, в-ва из окр среды + эффекторы-фитогормоны, в-ва из внешней среды 4 Генная регуляция позволяет кл-ке работать в режиме экономии белков не нужных в данный момент (Жакоб, Моно модель). Все виды внутриклеточной регуляции влияют др на др: генетич-мембр: наследств. Инф о белках мембран в геноме; мембр-генетич: ядерная мембр регулирует выход иРНК; мембр-ферм: через мембр поступ-т в-ва для построения мол-л ферм; ферм-мембр: ферм встроены в мембр, синтез компонентов мембр; ферм-генетич: биосинтез белка при помощи ферм; генетич-ферм: информ о ферм в геноме.
22. Фотосинтез – это процесс преобразования энергии солнечного света в энергию хим св органических соединений, синтезируемых из углек газа и воды. К фотосинтезу способны некоторые цианеи (прокариоты), зел растения. В ходе эволюции идет совершенствование процесса фотосинтеза: у бактерий нет спец органоидов и фотосинтетич функцию выпол выросты мембраны. У водорослей – хромотофор, у зел раст – хлоропласты (гранальный и агранальный). Чтобы свет оказал свое действие он должен быть поглощен фотосинтетич пигментами. В процессе эволюции сформиров след пигменты: бактериохлорофиллы у бактерий; хлорофиллы А и В, фикобелины и каратиноиды у раст. У низших все, у высш только хлорофилл и каротиноиды. Все молекулы пигментов вмонтированы в мембраны.
В процессе фотосинтеза выделяют : световую и темновую (метаболизм углерода, т.к свет активизир ферменты) фазы. Они разделены в пространстве: световая в мембране тилакоидов, темновая в строме хлоропласта.
СВЕТОВАЯ ФАЗА: делится на фотофиз и фотохим этапы. ФОТОФИЗ: молекулы пигментов образуют светособирающие антенны. Энергия со всех ССА стекается к молекуле-ловушке. Она вместе с пигментами ССА образует фотосинтетическую единицу. Молекула-ловушка является частью реакционного центра. Реакционный центр состоит из первичного донора электронов и первичного акцептора. Реакционный центр и фотосинтетич единица обр-т фотосистему. Выделяют два типа фотосистем: ФС 1 более древняя (аноксигенный фотосинтез, бактерии) и ФС 2 (если обе-оксигенный). Т.о. фотофиз этап закл в том что кванты света поглощ молекулами пигментов, энергия передается на молекулу-ловушку и в РЦ идет первичное разделение зарядов. ФОТОХИМ: фотохим р-ция – это р-ции, при кот фотоэнергия преобраз в энергию органич соединений, в первую очередь АТФ-фотосинтетическое фосфолиривание или фотофосфорилирование. Электрон от реакционного центра далее передается по электронтранспортной цепи (ЭТЦ). ЭТЦ представляет систему молекул переносчиков располож в строго определенном порядке в соответствии со своим окислительно-восстан потенциалом выраж в вольтах. Транспорт может быть циклическим (более простой и древний) и нециклич. ЦИКЛИЧ- участвует одна ФС1. И первичным донором и первичным акцептором одного и того же электрона яв молекула Р700, продукт- АТФ (у бактерий, аноксигенный фотосинтез). НЕЦИКЛИЧ: участвуют ФС1 и ФС2. Первичным донором служит молекула ловушка ФС2, а акцептором НАДФ+. Фосфорилирование происходит на пути транспорта электрона от ФС1 к ФС2. Продукты-кислород (фотолиз воды), восстан кофермент НАДФ*2Н и АТФ (фосфорилирование). Механизм фосфорилирования - хесмоосмотическая гипотеза Митчелла (при работе ЭТЦ мембрана поляризуется возникает µH , который используется для синтеза АТФ). Т.о продукты световой фазы: АТФ, восстан кофермент НАДФ*2Н, кислород.
МЕТАБОЛИЗМ УГЛЕРОДА: Происходит восстановление СО2 до углеводов за счет прод свет фазы (АТФ, НАДФ*2Н). Эволюциооно более древним является С-3 тип фотосинтеза -назван по первому устойчивому продукту-фосфоглицериновая к-та(ФГК) (в основе цикл Кальвина), но возник и др типы для адаптации у условиям: С-4 у троп и субтроп раст ( кукуруза, просо, сорго). Первый продукт- щавелево-уксусная к-та. САМ – фотосинтез у суккулентов( толстянка, кактус). Устьица этих раст днем закрыты, поэтому первичная фиксация СО2 идет ночью при откр устьицах. Процесс фотосин-за разделен во времени, а у С-4 в пространстве (карбоксилирование 2 раза: в кл-ках мезофилла и в кл-ах обкладки провод-го пучка).Цикл Кальвина этапы: карбоксилирование (рибулезодифосфат-карбоксилаза (РДФ)-первичный акцептор), восстановление (присоед-ие Н+ от НАДФ*2Н, энергия АТФ), регенерация (перестройка из 5 триоз в 3 пентозы-пополнение первичного акцептора СО2-РДФ), синтез углеводов (для получения настоящего углевода цикл должен пройти 4 раза).
Механизм фотофосфорилирования АДФ объясняет теория П. Митчелла. Согласно этой теории, трансмембранный перенос пластохинонами электронов и протонов в одну сторону чередуется с переносом цитохромной системой в обратную сторону электронов. Поэтому по одну сторону мембраны накапливается избыток протонов и возникает электрохимический мембранный потенциал. Его энергия используется для синтеза АТФ при разрядке мембраны в результате транспорта протонов через мембрану посредством Н+-АТФазы, которая действует как АТФсинтетаза. Этот фермент выглядит как грибовидная частица на поверхности мембраны тилакоидов. Его молекула состоит из двух частей: головки, выступающей с наружной стороны мембраны и содержащей активный центр фермента, и ножки, погруженной в мембрану и представляющей канал, через который передвигаются протоны.
Фотосинтез зависит от условий окруж среды: света – для каждой эко группы свое значение-мезофиты 20 тыс люкс; тененевыносливые- 2-4 тыс люкс. Температура: есть температур коэффициент Q10 – во сколько раз увеличив интенсив фотосин при увел темпер на 10 градусов. Вода – исходное вещество. Концентрация кислорода – конеч продукт. При его высоком содержании тормозится фотосинтез. Наблюд-ся явл-е фотодыхания-светозависимое поглощение О2 и выделение СО2 с использ-м в качестве субстрата промеж прод цикла Кальвина (двойственная природа РДФкарбоксилазы-может присоед-ть СО2 и О2) Минеральное питание – для процесса фотосин необходимы P N Fe Mg K S Cu. ЗНАЧЕНИЕ: 1. образование и накопление органич массы, ежегодно 160-200 млрд тонн. 2. выделение кислорода 100-150 млрд тонн – раст участ в образ озонового экрана; 3. поддержание конц углек газа в атмосфере на определ уровне за счет поглощ 250-400 млрд тонн- сниж парникового эффекта.
23. Способы получения энергии. Обязательным условием существования любого организма является постоянный приток питательных веществ и постоянное выделение конечных продуктов хим р-ций, происходящих в кл-ках. Совокупность хим р-ций, происходящих в организме, называется обменом веществ или метаболизмом. В зависимости от общей направленности процессов выделяют катаболизм и анаболизм. Катаболизм (диссимиляция)—совокупность реакций, приводящих к образ-нию простых соединений из более сложных (р-ции гидролиза полимеров до мономеров и расщепление последних до углекислого газа, воды, аммиака, т. е. реакции энергетич обмена, в ходе которого происходит окисление орг вещ-в и синтез АТФ. Анаболизм (ассимиляция) — совокупность реакций синтеза сложных орг вещ-в из более простых (фиксация азота и биосинтез белка, синтез углеводов из углекислого газа и воды в ходе фотосинтеза, синтез полисахаридов, липидов, нуклеотидов, ДНК, РНК и др вещ-в). Синтез веществ в кл-ах живых орг часто обозначают понятием пластический обмеи, а расщепление веществ и их окисление, сопровождающееся синтезом АТФ, —энергетическим обменом. Оба вида обмена составляют основу жизнедеятельности любой клетки, а следовательно, и любого организма и тесно связаны между собой. С одной стороны, все реакции пластического обмена нуждаются в затрате энергии. С другой стороны, для осуществления реакций энергетического обмена необходим постоянный синтез ферментов, так как продолжительность их жизни невелика. Кроме того, вещ-ва, используемые для дыхания, образуются в ходе пластического обмена (например, в процессе фотосинтеза). Существует 2 способа получения энергии у ж.о.-брожение и дыхание. Брожение-анаэробный окислительный распад сложных органических соединений до простых орг. и неорг. с выделением энергии. Характерен для м.о. и в экстримальных условиях для раст. Существует несколько типов брожения (по продукту): молочно-кислое, эволюц-нно более древнее, вызвано молочно-кислыми бактер-ми (молочно-кислый стрептококк, молочная, болгарская палочки). Наиб известно спиртовое (дрожжи). Брожение эволюционно более древнее, т к атмосфера Земли носила восстановительный характер. После появления фотосинтеза и накопления в атмосф кислорода появляется новый способ получения энергии-дыхание. Дыхание-аэробный окислительный распад слож орг вещ-в до простых неорг с выделением энергии. Оно характерно для растений, м. о. животных. В качестве субстрата м б любые орг в-ва, способные к окислению (белки, жиры, углеводы). Существует несколько путей дыхательного обмена: дихотомический (гликолитический), апотомический (пентозофосфатный), глиоксилатный. Наиб распространен дихотомический Этапы: 1) Подготовительный (у раст, м.о-в цитоплазме; у животных, чел-в пищеварит. сист) Происходит гидролиз сложных орг вещ-в до простых: белки-аминокислоты, жиры-глицерин и жирные кисл, углеводы-глюкоза. 2) неполное окисление (у всех в цитоплазме) глюк до ПВК (чаще всего гликолиз), аминокисл-окислительное дезаминирование, жиры-окислительное декарбоксилирование. На этом этапе происх-т субстратное фосфорилирование за счет макроэргич связи в мол-ле 1,3 диФГК, обр-ся 2 АТФ. 3) Кислородный (полное окисление) происходит в митохондриях делится на фазы: а) окислительное декарбоксилирование ПВК б) Цикл Кребса в) ЭТЦ дыхания первая и втор в матриксе, третья в мембр-х крист. Конечным акцептором 2е и 2Н+ явл-ся О2 обр-ся Н2О. Синтез мол-л АТФ осущ-ся через грибовидные тела, наход-ся в мембране митохондр. Шляпка грибовидного тела располож на внутренней стороне мембраны. Синтез АТФ на мембр крист-окислительное фосфорилирование. При выбросе каждой пары Н+ синтезир-ся 1 АТФ. Энергетический выход пути-38 мол-л АТФ на 1 моль глюкозы. КПД 56%. Остальное рассеивается в виде тепла. Апотомический путь: энергитич выход почти такой же, его главное значение-участие в пластическом обмене: синтез-ся углеводы с разл длиной цепочки в т ч пентозы, включ-ся в синтез нуклеотидов. Глиоксилатный путь: у бактерий и плесневых грибов, в семенах масличных растений, в стареющих листьях, у животных отсутствует. Дых. субстрат-липиды. Происх-т отщепление 2-х углеродных фрагментов, из которых обр-ся мол-лы Ацетил Ко-А. Предполагают, что гены, кодирующие ферм этого цикла есть у всех раст, но экспрессируются в стареющих листьях, семенах.
Значение дыхания: 1 получение энергии; 2 биосинтетическая вода; 3 обр-ся промежуточные метаболиты, которые могут выходить из энергитич обмена и включаться в др ветви обмена вещ-в.
Дыхание более энергитич выгодно-38 АТФ, брожение-2 АТФ. Существует связь между дыханием и брожением: 1) в кл-ке раст сущ-т информ о всех ферм брожения. 2) Эффект Пастера-переключение с дыхания на брожение в определенных условиях (отсутствии кислорода). Первые 2 этапа в дыхании и брожении одинаковы, метаболическая вилка нах-ся на уровне ПВК. При брожении увеличив-ся скорость разложения субстрата, этиловый спирт ядовит, поэтому долго растение не может исп-ть брожение. Регуляция энергетич обмена: 1) Переключение с одного способа получения энергии на др 2) переключение с одного пути дых обмена на др. метаболич вилка на уровне глюкоза-6фосфат 3). Изменение скорости энергитич обмена: а)субстратный контроль (качество-белки, углеводы, липиды); кол-во; доступность (углеводы-крахмал, глюкоза) б) на уровне ферментов: 1. через аллостерич эффект: АТФ/АДФ; НАД/НАД 2Н 2. Через изменение физ-хим св-в цитоплазмы: сдвиг рН, оводнение, наличие ионов; г) генная регуляция связана с новообраз-м ферментов. В растит орг-ме обр-ся много промеж метаболитов, которые могут включаться в разл ветви обмена вещ-в. Самая тесная связь с фотосинтезом. Глюкоза может откладыв-ся в виде крахмала, может включаться в компонент кл стенок, может окисляться при энергитич обмене. Пересечение с белковым обменом осущ-ся в рез-те аминирования ряда орг кислот. Большая метаболич вилка наход-ся на уровне Ацетил-Ко А, здесь возможно пересечение с липидным обменом через образ-ие жирных кислот и с гормональным обменом. Пентозо-фосфатный путь с нуклеиновым обменом и гормональным (поставщик пентоз). Цикл Кребса-«метаболич котел кл-ки».
24. Физиологические основы устойчивости. Погодно-климатические условия на Земле становятся менее благоприятными поэтому все ж. о. должны приспосаблив-ся к изменяющимся условиям среды. Адаптация- процесс приспособления строения и функций организма к условиям среды. Достигается с помощью различных механизмов: генетических, биохимических, физиологических, морфоанатомических и др. Живые системы обладают свойством сочетать устойчивость, т. е. относительную стабильность, при изменяющихся условиях среды (гомеостаз) и подвижность, т. е. приспособление к этим условиям. Под действием различных неблагоприятных факторов в ж. о, как правило, развивается особое состояние, называемое стрессом. Теория стресса сформулирована Гансом Селье 1982, и термин «стресс» (от англ. stress — напряжение). Внешние факторы, действующие на биологическую систему и вызывающие стресс- стрессоры, а стресс- состояние организма, формирующееся в ответ на их воздействие. Например, «температурный стрессор», «осмотический стрессор» и т. д. Реакции на стрессорные воздействия лишь при некоторых условиях являются патологическими, в принципе же они имеют адаптивное значение, и поэтому были названы Селье «общим адаптационным синдромом» (употреблял как синонимы). Обычно выделяют три фазы реакции организма на воздействие неблагоприятных факторов: первичная стрессовая реакции (по Селье: тревоги), адаптации (по Селье: резистентности) и истощения. 1Фаза: наблюдаются значительные отклонения в физиолого-биохимических процессах, проявляются как симптомы повреждения, так и защитная реакция. Если воздействие слишком велико, организм погибает еще в стадии тревоги в течение первых часов. Если этого не случилось, реакция переходит во вторую фазу. 2 Фаза: организм либо адаптируется к новым условиям существования, либо повреждения усиливаются. При медленном развитии неблагоприятных условий организм легче приспосабливается к ним. После окончания фазы адаптации организм нормально функционир-т в неблагоприятных условиях уже в адаптированном состоянии при общем пониженном уровне процессов. 3 Фаза повреждения (истощения, гибели)- усиливаются гидролитические процессы, подавляются энергообразующие и синтетические реакции, нарушается гомеостаз. При сильной напряженности стресса, превышающей пороговое для организма значение, организм гибнет. Адаптационный процесс (адаптация в широком смысле) протекает постоянно и осуществляет «настройку» организма изменениям внешней среды в пределах естественных колебаний факторов. Реакция организмов на изменившиеся условия является комплексной (изменения биохимических и физиологических процессов). Эти изменения могут быть: неспецифич-однотипные реакции организма на действие разнородных стрессоров или разных организмов на один и тот же стресс-фактор; специфические- ответные реакции, качественно отличающиеся в зависимости от фактора и генотипа. Важнейшей неспецифич реакцией кл-ок на действие стрессоров-синтез особых белков (общие и специальные). Ряд таких белков идентифицирован в 80—90-е годы. Установлены гены, кодирующие белки и показано, что стресс индуцирует экспрессию целого ряда генов.. Стрессовые белки синтезируются в ж.о. в ответ на различные воздействия: анаэробиоз, повышенные и пониженные температуры, обезвоживание, высокие концентрации соли, действие тяжелых металлов, вредителей, а также при раневых эффектах и ультрафиолетовой радиации. Стрессовые белки разнообразны и образуют группы высокомолекулярных и низкомолекулярных белков. В результате изменения действия фактора или факторов происходит переключение жизни клетки на стрессовую программу, одновременно на многих уровнях регуляции. Тормозится экспрессия генов, активность которых характерна для жизни клетки в нормальных условиях, и активируются гены стрессового ответа. Активирование генов стресса: стресс- сигнал -> рецептор в плазмалемме -> сигнальная цепь в цитозоле (протеинкиназы, фосфотазы) —> транскрипционный фактор в ядре —> промотор стресс-индуцированного гена -> мРНК -> белок -> защитная роль в растении. В настоящее время исследованы промоторы различных стресс-индуцируемых генов (сегмент из 6 нуклеотидов активируется АБК, сегмент из 9 нуклеотидов активируется осмотическим стрессом) Ответ на стресс развивается достаточно быстро через 5 мин появляются мРНК, кодирующие стрессовые белки, уже через 15 мин в клетках обнаруживаются стрессовые белки. Их синтез постепенно нарастает, достигая максимума, а затем ослабевает. После окончания воздействия синтез стрессовых белков прекращается и возобновляется синтез белков, характерных для клетки в нормальных условиях. Важнейшей неспецифической реакцией на неблагоприятные воздействия является изменение свойств мембран, что связано с перестройками в их структуре (сдвиги в соотношении степень ненасыщенности различных групп жирных кислот). Большую роль играет Са2+ в поддержании структуры хроматина, в регуляции активности ферментов в митохондриях и хлоропластах. Изменение концентрации кальция запускает специфические мембранные каналы и транспортные системы, а также вызывает структурные изменения в клетке. Нарушение структуры мембран приводит к многочисленным изменениям в метаболизме. Возникает недостаток энергетических ресурсов, происходит временная активацией дыхания. Возрастание активности гидролитических процессов что ведет и к накоплению различных протекторных соединений (пролин обр-т гидрофильные коллоиды, что удерживает воду и защищает белки от денатурации; полиамины - увеличивают стабильность РНК, ДНК, рибосом, стабилизируют мембраны). Существенную роль в ответе организмов на стрессоры играет гормональная система (возрастает количество абсцизовой кислоты, этилена изменяется соотношение фитогормонов урастений, соотношение гормонов у животных). Высказывается мнение, что у растений в отличие от животных при неблагоприятных условиях ведущую роль играют гормоны, тормозящие их функциональную активность. Это обеспечивает организму торможение роста и вхождение в покоящееся состояние. При действии неблагоприятных факторов важным для растения, является сохранение нормальных донорно-акцепторных отношений, поскольку при снижении запроса на ассимиляты, уменьшается фотосинтез. Устойчивость растений против неблагоприятных условий имеет разный характер: избегание воздействия (суккуленты, эфемеры); устойчивость- способность в процессе адаптации перестраивать как скорость, так и направление метаболических реакций таким образом, чтобы и в изменившихся условиях среды вырабатывать все необходимые соединения. Разный уровень устойчивости обусловлен биологическими особенностями видов. Для растений, устойчивых к действию стрессоров, показана большая стабильность клеточных мембран по сравнению с неустойчивыми (мембраны холодостойких растений отличаются большим содержанием ненасыщенных жирных кислот (позволяет сохранять жидкостное состояние при пониженных температурах)). Существуют различные виды устойчивости в зависимости от действующего стрессора (водный дефицит, высокие температуры, холодостойкость, морозостойкость, солеустойчивость). Животные, человек: Одним из механизмов приспособления к окружающей среде является поведение животных, которое можно назвать эффективным средством для поддержания гомеостаза в организме. Поведенческие акты являются первыми показателями взаимосвязи организма и среды, по которым можно судить о характере комфортности условий. На их формирование оказывает влияние нервная система организма. Одним из многих тестов, характеризующих естественную устойчивость организма, является состояние высшей нервной деятельности (ВНД) человека и животных. И. П. Павлов представил классификацию типов нервной системы. В ее основу были положены три основных свойства нервных процессов: сила, уравновешенность и подвижность процессов возбуждения и торможения (безудержный, сильный уравновешенный, слабый). Чем чувствительнее организм к колебаниям внешних условий, тем значительнее изменяются его клинико-физиологические показатели. В процессе эволюции организмы смогли выработать ряд путей нейтрализации действия неблагоприятных факторов. Активный путь — развитие сопротивляемости, т. е. регуляторных процессов, которые позволяют осуществить все функции организма, несмотря на неблагоприятные факторы (теплокровные животные — птицы и млекоп-ие -поддержание постоянной температуры, опт для биохимических процессов в кл-ах тела, верблюд обеспечивает потребности во влаге биохимич окислением собственного жира.). Это требует больших затрат энергии, которую им надо постоянно восполнять. Это -т. н. физиологические адаптации. Поведенческие адаптации- создание убежищ, передвижение с целью выбора оптимальных температурных условий, особенно в условиях экстремальных температур. Приспособительное поведение может проявляться у хищников в процессе выслеживания и преследования добычи, а у жертв — в определенных ответных реакциях (например затаивание). Обычный для животных способ приспособления к неблагоприятным периодам — миграция (сайгаки на зиму в малоснежные южные полупустыни; летом в более влажные северные степи). Активное сопротивление влиянию неблагоприятной среды способствует развитию морфологической адаптации отдельных видов, т. е. приобретению таких особенностей внешнего строения, которые обеспечивают выживание в экстремальных и успешную жизнедеятельность организмов в привычных для них условиях (у китообразных приспособления к скоростному плаванию, особая структура кожи у акул). Морфологический тип приспособления животного или растения- жизненная форма вида. При этом разные виды могут иметь сходную жизненную форму, если ведут близкий образ жизни. Примерами здесь могут быть кит (млекопитающее), пингвин (птица) и акула (рыба). Пассивный путь -подчинение жизненных функций организма изменению факторов среды (недостатке тепла - понижение уровня метаболизма- экономное использованию энергетических запасов). При резком ухудшении условий среды организмы разных видов могут практически приостанавливать свою жизнедеятельность и переходить в состояние т. н. скрытой жизни. Переход в состояние глубокого анабиоза, при котором резко замедляется обмен веществ, расширяет возможности выживания организмов в самых экстремальных условиях (высушенные семена и споры растений, коловратки, нематоды могут выдержать температуры ниже — 200°С; Некоторые бактерии, годы в неактивном состоянии - благоприятные условия -«пробуждения». Явление временного физиологического покоя в индивидуальном развитии некоторых животных и растений, обусловленного неблагоприятными факторами внешней среды, называется диапаузой. В геноме живых организмов заложен богатый арсенал программ, которые способствуют его выживанию при изменении условий окружающей среды. Некоторые программы активизируются только при изменении условий и в обычных условиях ничем себя не проявляют. Другие находятся в наличии у незначительного числа особей. При влиянии неблагоприятных или мутагенных факторов, когда большинство популяции погибает, особи с набором устойчивых генов благополучно выживают и дают начало новой популяции, состоящей из их потомков. У каждого рода организмов устойчивость различна. Высокая генетическая устойчивость организмов говорит о богатстве их генофонда. Хорошую генетическую устойчивость демонстрируют бактерии, грызуны, насекомые, злаковые растения.
25. Мех-мы регуляции и интеграции физиолог проц-ов в растит орг-ме. Живые организмы обладают определенной структурной организацией (одноклеточной, многокл). Регуляция обеспечивает гомеостаз организма, сохранение параметров внутренней среды и создание условий для развития растений. Т.к. первые ж. о. были одноклеточными, то первыми в ходе эволюции возникли внутриклеточные системы регуляции (мембранная, ферментативная, генетическая). Мембранная регуляция осущ-тся путем сдвигов в мембранном транспорте, связыванию или освобождению ферментов и регуляторных белков, изменением активности мембранных ферментов. У эукариот с появлением ядерной обол-ки роль мембр регуляции возросла. Мембр. Регуляция контактная (ЭПР, АГ, где происх-т достройка и модификация секреторных белков, связывание мембр-ми освобожд-ся ферментов меняет их активность). Дистанционная (путем избирательной доставки ферментов, ионов, субстратов, удаление продуктов). Генетическая регуляция осущ-ся на уровне репликации и экспрессии генов (биосинтез белка): на уровне транскрибции и посттранскриционном уровне (процессинг, трансляция, созревание белка). Молекулярные механизмы осущ-ся за счет рН, активности ферментов, ионов. Не все гены в организме постоянно находятся в рабочем состоянии, существует сложная система регуляции. 1 гипотеза: 1961 Жакоб, Моно модель генетического контроля синтеза белков для прокариотической клетки. 2 гипотеза: 1997 Шарп и Робертс – гены эукариот это не непрерывные белоккодирующие нуклеотидн. последовательности, а мозаика из экзонов (кодирующие белки) и интронов (не кодирующие, несут регуляторную функцию). В геноме кроме структурных генов есть гены регуляторы, несущие информацию о белках регуляторах (их ф-ция включение/выключение гена). Белки регуляторы: 1 репрессоры (присоединяется к оператору не дает осущ-ть транскрибцию); 2 активаторы (способствуют транскипции). На способность репрессора соединяться с оператором влияют некоторые низкомолек-е в-ва (гормоны, аминокисл) –эффекторы. 2 вида: 1 корепрессор соед-ся с неактивной формой репрессора, переводит в активную, блокирует транскрипцию; 2индуктор соед-сь с репрессором мешают его способности взаимодействовать с оператором. Внутриклеточная регуляция осущ-ся так же за счет таких св-в цитоплазмы как вязкость, подвижность, эластичность. Ферментативная регуляция. Ферменты-специфич белки, присутствующие во всех живых клетках, играющие роль биокатализаторов в р-циях обмена вещ-в. Регуляция работы ферментов. Ферм м б конститутивными (постоянно присутствуют) и индуцированными (появл-ся иногда). 2 пути регулир-я активности ферм: 1) Мех-мы направл-е на поддержание клет-го гомеостаза 2) Мех-мы слежения за сигналами, поступ-ми в кл-ку. Они реализ-ся через след. Мех-мы: 1. Превращение неактивных предшеств-ов ферм (зимогенов) в активные ферменты; 2 Изостерический эф-т для низкомолек ферм, основан на конкурентном ингибир-ии (определенные в-ва конкурируют с субстратом за акт центр ферм) 3Аллостерический эф-т характ-н для крупных мол-л, имеющих дополнит сайт связывания с эффектором-аллостерический центр. – эффекторы-некот фитогормоны, конечные прод р-ции, в-ва из окр среды + эффекторы-фитогормоны, в-ва из внешней среды 4 Генная регуляция позволяет кл-ке работать в режиме экономии белков не нужных в данный момент (Жакоб, Моно модель). Все виды внутриклеточной регуляции влияют др на др: генетич-мембр: наследств. Инф о белках мембран в геноме; мембр-генетич: ядерная мембр регулирует выход и РНК; мембр-ферм: через мембр поступ-т в-ва для построения мол-л ферм; ферм-мембр: ферм встроены в мембр, синтез компонентов мембр; ферм-генетич: биосинтез белка при помощи ферм; генетич-ферм: информ о ферм в геноме.
С появлением многоклет-ти появляются межклеточные и оргпнизменные системы регуляции. Межклеточные-трофическая: некоторые органы растения (корень, части цветка, плоды) нуждаются в готовых орг вещ-вах, получаемых от фотосинтезир-х частей. Надземная часть нуждается в поступлении воды и минеральных вещ-в из корневой системы. Если раст испытывает минер голод то развитие происходит, но изменяются размеры, продуктивность (пасынкование вершкование). Электрофизиологическая: у раст сущ-т электрофизиологич явления: 1. Разность потенциалов между различными частями растения (избират прониц-ть мембр, перераспред-ие ионов). 2. Распространяющийся потенциал действия (между разл участками возникает ток 0,1-0,4 микроА; под действием раздражителя деполяриз-ия, затем реполяриз-ия). ПД у венериной мухоловки 25 см/с, у обычных раст-0,08-0,5 см/с. ПД распростр-ся по плазмолемме и плазмодесмам, паренхимным кл-кам флоэмы и протоксилемы проводящих пучков. Если подействовать на побег-импульс в корень(усиление подачи минер вещ-в вверх) и наоборот. Гормональная: фитогормоны-продукты естественного обмена вещ-в, которые образуются в одних тканях и органах растительного орг-ма и в малых кол-вах оказывают регуляторное влияние на физиологич процессы к п в др тканях и органах. Делят на стимулирующие (ауксины-верхушка побега, апикальное доминирование; гиббереллины-в листьях, переход к цветению, удлинение стебля; цитокинины-апекс корня, стимулир-е деления, апикальное доминирование; брассины-увеличивают длину и толщину междоузлий) и ингибирующие (абсцизины-хлоропласты, торможение роста; этилен-стареющие листья, плоды, увелич роста в толщину, уменьш в высоту; вещ-ва фенольной природы). Важно соотношение фитогормонов: после прорастания доминируют стимулир-ие, после цветения-ингибирующие. Метаболические вилки Ацетил-Ко-А может быть предшественником для ауксина (если мало азота) и для цитокининов, гиббереллинов, абсцизовой к-ты (если достаточно). Организменные системы регуляции: доминирующие центры-у растений имеются два полюса-верхушка побега и кончик корня. Зоны ткане- и органообразования, аттрагирующие зоны и зоны синтеза важнейших фитогормонов-ауксина и цитокенина. Наличие доминирующих центров приводит к созданию физиологических градиентов: осмотического давления, концентрации фитогормонов, рН. Что обеспечивает полярность у растения. Связь осуществляетсяпри помощи канализированной связи-проводящая ткань (передача пит вещ-в, гормонов, ПД). Временная интеграция осущ-ся системой взаимосвязанных осцилляций (ритмов). Могут быть анатомические (закономерное чередование листьев, узлов, пазушных почек) и физиологические (сезонные-листопад, интенсивный рост; Циркадные(суточные) цветки, листья) осцилляции. Регуляторные контуры: межклеточные системы функц-т совместно в виде регуляторных контуров с образованием +и – связей, обеспечивают мех-мы раздражимости. Рецептором восприним-ся стимул, сигнал в электриич. или гормональной форме передается по каналам связи, достигает компетентного органа, индуцирует его активность и осущ-ся ответ на действие раздражителя.
26. Биотехнология. Термин-К. Эрикл 1917г. Биотехнология как наука рассматривается в двух измерениях: классическом–наука о методах и технологиях производства, хранения переработки различных вещ-в с использованием обычных растений, животных, м о (не трансгенных);
современном–наука о генных, инженерных и клеточных методах создания и использования генетически модифицированных животных, растений, м о (трансгенных, рекомбинантных, химерных). Она делится на: IГенная инженерия- выделение нужного гена из генома одного организма и введение в геном др орг-ма (трансгенные оргн-мы) или выбиванием гена (нокаутные орг-мы). Основана: 1972 Поль Берг синтезир-на рекомбинантная мол-ла ДНК. Задачи: 1.Получение раст, устойчивых к насекомым; 2.Улучшение качества пищевых продуктов; 3.Улучшение товарных качеств декоративных раст; 4.Повышение устойчивости к засухе, заморозкам; 5.Биосинтез инсулина, антибиотиков. II.Хромосомная инженерия–введение в генотип пары чужих гомологичных хромосом или замещение одной пары на др. (снижение или увелич числа хромосом). Задачи: получение полиплоидов, гаплоидов, анеуплоидов. III.Клеточная инженерия- культивирование отдельных кл-ок или групп кл-ок на питательных средах, где они обр-т клет культуры регенерирующие во взрослые раст. Возникла в 50-е годы 20 века, когда доказали свойства тотипотентности (1 кл-ка раст-один организм). Задачи: 1.Быстрое получение биомассы раст кл-ок в том числе необходимых для синтеза втор метаболитов 2.Оздоровление раст-выделение меристемы и ее культивирование in vitro. 3.Клональное размножение in vitro. 4.Отдаленная гибридизация. 5.Соматическая гибридизация-выделение протопластов и их слияние. Каждому направлению хар-ны свои методы. Клеточная инженерия: культивирование в жидкой пит среде (глубинное культивир-е). Суспензионная культура-выращивание отдельных кл-ок или небольших групп во взвешенном состоянии в жидкой пит среде на аппаратах роллерного типа (качалки, шейкеры, роллеры). Скорость- 100 об/мин, выше-механич разрушение, ниже не хватает воздуха. 1 способ из каллюсных тканей (разрыхл-т пектиназой, удаляют Cа+): Колба V 500-1000 мл, суспензии 1/5-1/10 от колбы, кол-во каллюса 2-5 г. 2 способ из экспланта по Стюарту. Для раст, у которых не обр-ся каллюс на тв пит среде. Эксплант в жидк пит среду, на качалку, кл-ки отрываются и переходят в суспензию, фильтруют. Пассеруют через каждые 14 дней. Плотность кл-ок 107-108, мах-1012, больше-не хватает О2, меньше-не хватает кондиционирующего фактора (метаболит, контролирующий очередность деления). Кривая роста S-образная. Назначение суспензий: а)получение одиночных кл-ок для генетических экспериментов(генетич неоднородность, изменение плоидн-ти), б)получение протопластов (для получения гаплоидов) в)выращивание кл-ой биомассы, главное-г) получение вторичных метаболитов. Первичн метаболиты-соед-я обра-ся во всех жо и служащих для построения макромолекул ферментов, клет структур (Белки, жиры, углеводы, нк). Втор-низкомолекулярные соед-я, не являющ-ся обязательными для роста и функц-я кл-ки (алкалоиды, терпеноиды, фенолы). Используются для получения лекарств, красителей, вкус добавок, косметики. Мак снотворный-кодеин, опиум. Преимущ-ва получения: Получение круглый год, использование раст не произрастающих в наших условиях, нет необходимости ждать пока растение вырастет, занимают набольшие площади, превышает продуктивность в обычн условиях, автоматизация процесса. Культура изолированных кл-ок и тканей в селекции растений. I Преодоление прогамной несовместимости в том случае, когда невозможно оплодотворение между раст-ми в нормальных условиях. Причины: физиологические-не соответствие во времени созревания пыльцы и завязи; не соответствие секрета рыльца матер-го и пыльцы отцовского; морфологические-короткая пыльцевая трубка, блокирование роста пыльцевой трубки. Способы оплодотворения in vitro: 1Культивирование на пит среде завязи, на которую наносится готовая пыльца; 2Завязь вскрывается и на пит среде культивир-т семяпочки, а рядом-пыльцу. II Преодоление постгамной несовместимости (возникают при отдаленной гибридизации после оплодотв-я, в результате которого обр-ся невсхожие семена). Причины: расхождение во времени развития зародыша и эндосперма; слабое развитие эндосперма; нарушение диффер-ки зародыша. Зародыш изолируют и культивируют на пит среде-эмбриокультура (ржаные, пшенично-колосняковые гибриды). III Получение гаплоидов in vitro. Их роль: проявление действия гена, определяющего рецессивный признак; проявление мутаций (к п рецессивны); выяснение влияния условий на характер проявления гена. Способы получения гаплоидов: 1Андрогенез-из изолорованных пыльников и микроспор.2Гиногенез-из изолированных семяпочек 3Псевдогамия-из зародыша, у которого из-за несовместимости хромосом были потеряны отцовские хромосомы.4Селективная элиминация-при скрещивании культурного и дикорастущего вида происх-т утрата хромосом дикого вида. Селекция раст-й на клеточном уровне (использ-т селективные факторы соли, гербициды): 1 Прямая-обрабат-т фактором высокой концентрации и отбирают выжившие; 2Непрямая-избирательная гибель метаболически активных кл-ок и медленное приспособление к фактору кл-ок, наход-ся в покое. 3Визуальная-кл-ки отбирают по анатомическим или биохимическим признакам.
Генетическая инженерия- ветвь молекул генетики, которая конструирует генетич системы вне орг-ма с последующим введением их в живую кл-ку. Rek ДНК-фрагмент чужеродной ДНК, сод-йе важные св-ва (гены) и вектор-ДНК, обеспечивающий репликацию. Этапы: 1Выделение ДНК; 2Ращепление в специфич участках (ферм-рестриктазы); 3Секвенирование-опред-ие последовательности нуклеотидов, определение границ гена; 4Литрование-сшивание с помощью ДНК-лигазы; 5Введение в живую кл-ку с помощью вектора (вирусы для животных-герпеса; для раст-ий-табачной мозаики; агробактерии-бактериальные плазмиды). Бактерии, дрожжи-получение инсулина, соматотропина, интерферона. Растения-улучшение аминокисл-го состава, устойчивость к патогенам, продуценты гормональных препаратов. Животные-повышение устойчивости, улучшение состава молока, секркция чел белков, повышение продуктивности, доноры органов и тканей для чел (островки Лангерганса в печени свиньи).
27. Водоросли. По соврем классифик-ии делят на 10 отделов: 1)сине-зеленые (анабена), 2)пирофитовые (криптомонада) 3)золотистые (динобрион), 4)бурые (ламинария), 5)красные (лемания), 6)харовые (хара), 7)диатомовые (меридион), 8)желто-зеленые (вошерия), 9)евгленовые, 10)зеленые (кладофора, улотрикс). Водоросли-первичные фототрофные орг-мы. С ними связано появление фотосинтеза. Строение-одноклет и многоклет, сифональная структура (таллом не поделен на клетки, но в цикле есть однокл стадии). Однокл м б шаро-, веретено-, нитевидными. Наим-сине-зел, наиб-харовые. Оболочки: протопласт окружен плазмолеммой (в отличие от высших), тека-наиболее плотная (вольвоксовые, хризомонады), пектиновая, целлюлозно-пектиновая (наиб удачна-опора,рост, защита, проницаемость). Наиболее толстые обол-ки у покоящихся стадий, наименее-у гамет. Обол-ки часто пропит-ся орг в-вами-кутин, лигнин. Все кроме сине-зел эукариоты. В строении присутствуют-ядерн сок и хроматин-е структуры, цитоплазма и органеллы: митохондрии, диктиосомы, ЭПС, микротрубочки, лизосомы, вакуоли (пульсирующие-у подвижных, выведение воды и прод обмена в-в, хламидомонада; газовые у сине-зел-сниж объема, передвиж). Фотосинтетич аппарат: у сине-зел не оформлен, типичн- у зел (хлоропл, хлорофилл а, b, c, d); у кр, бурых-фикоксантин, фикобелин. Большое разнообразие хлоропл-ов по форме, размерам т д., пиреноид-место накопления крахмала (нет у высших и сине-зел). Стигма у зел, золотист, бурых в хлоропласте, у евгленовых рядом с двигательными органеллами. Все (кроме син-зел и красных) обр-т подвижные кл-ки в течение всей жизни или на опред этапе. Органы движ-реснички, жгутики, ложные реснички. Жгутики различные-равно-, разно-, гладкие и перистые. Водоросли дали начало наземным. Оч большое разнообразие в кл-ах и их компонентах-активный формообразов-ый процесс. Отдельные морфологич структуры: высшие раст имеют листостебельный тип строения, водоросли морфологич-ки разнообразны. Существуют основные морфологич структуры типа водоросли: 1) Амебоидная-для одноклеточных, лишенных тв клет обол-ки и пост формы тела (золотистые, желто-зел, пирофитовые); 2)монадная-однокл с тв кл-ой обол-ой (перипласт)-наличие 1-2 жгутиков, движение в воде (жел-зел, зел, золотист, пирофитовые), есть колониальные-собрание отдельных кл-ок монадного строения. У б-ва нет различий между кл-ми колонии, у некоторых-на вегетативные (более мелкие) и половые (более крупные). 3)коккоидная-отдельные кл-ки с тв обол-ой, в вегетативн состоянии лишены жгутиков и псевдоподий (образ-ие колоний со слизью и без), единственная для диатомовых; 4)пальмеллоидная-усложнение коккоидной, образ-ие крупных прикрепл-х к субстрату слизистых тел, содерж-их множество коккоидных кл-ок. В это состояние переходят некот однокл подв и неподвиж при неблагоприятн условиях; 5)нитчатая-талломы из различного кол-ва кл-ок в форме нити (простые, ветвящ-ся, прикрепл, свободн, колонии); 6)разнонитчатая-талломы дифер-ны на 2 части: стелящиеся и прямостоячие (золотист, бурые, зел, сине-зел); 7)пластинчатая-растут на субстрате прикрепляясь или распростершись (зел, бурые, кр); 8)сифональная-отсутствие кл-х прегородок и ядер (зел, жел-зел); 9)харофитная-крупное многокл слоевище линейно-членистого строения, состоит из главного побега с ветвями, внизу ризоиды, на побегах членистые мутовки с боковыми побегами (харовые). Размножение: вегетативное-деление однокл на 2 части; многокл-при механич разруш-ии слоевища. У харовых обр-ся клубеньки, перезимовывают и дают новые раст-я; нитчатые (улотрикс)-отд кл-ки округляются, накапливают пигменты, пит в-ва, утолщ-ся обол-ка-акинеты; у кр, бурых, харовых, зел есть стелящиеся побеги на которых могут вырастать новые слоевища. Бесполое: деление протопласта-выход подуктов из обол-ки материнск кл-ки-зооспоры и споры. Зооспоры в зооспорангиях. У сине-зел эндо и экзо споры, у б-ва эукариот размножение зооспорами. Виды спор водорослей: апланоспоры-зел, желт-зел, одеваются оболочкой внутри матер кл-ки; гипноспоры-могут нах-ся длительно в периоде покоя м б тетеро, би и моно;автоспоры-апланоспоры, приобретающие форму материнской. Выходят в воду через отверстие в спорангии, гаметы так же. Половое: образование зиготы. Неск-ко типов: 1слияние 2-х особей (однокл-ие)-хологамия; 2коньюгация (спирогира)-слияние 2-х вегетативных кл-ок. Различают типов пол-го процесса от размеров гамет: 1изогамия-равные кл-ки; 2гетерогамия-женская крупн, мужская-менее; 3оогамия-женская (яйцеклетка) без жгутиков крупнее, слив-ся со сперматозоидом; 4автогамия (диатомовые)-не увеличение числа особей, а их омоложение. Зиготы различны: могут сохранять жгутики и плавать в течение неск-х дней, недель. Органы поло-го и бесполого размнож-ия м б на разных особях-споры-спорофиты, гаметы-гаметофиты; м б одинакового строения. Для прорастания зиготы треб-ся комплекс оптим-х условий: темпер-ра, наличие биохим и биоактив в-в. Воспоизвед-ю и восстановл-ию водоросл спосбствует образ-ие цист (золот, жел-зел, диатомовые). При прораст-ии цист обр-ся одна особь, реже-несколько. Экологич группы: планктонные, бентосные, наземные, почвенные, горячих источников, снега и льда, соленых водоемов, в известковом субстрате. Значение: могут исп-ся в качестве индикаторов состояния водоем (биоиндикаторы), начальное звено в трофической цепи экосистемы водоема. Эволюция водорослей: Многие группы мало изменились со времени своего возникновения многие их виды сейчас вымерли. Крупных колебаний в видовом разнообразии и численности не было. Водные м о за млн лет менялись слабо, и современные формы водорослей существуют уже очень долго. Маловероятно, что какая-либо крупная группа водорослей появилась позже палеозоя или раннего мезозоя (240 млн.л н). Зеленые водоросли. Большинство - микроскопические пресноводные формы, многие-на почве. Бывают одно- и многоклеточными, образ-т нити, шаровидные колонии, листовидные структуры и т.д. Кл-ки подвиж (с двумя жгутиками) или неподвиж. Половое размножение - разных уровней сложности в зависимости от вида. Кл-ки содержат ядро и несколько четко оформленных хлоропластов. Род плеврококк-однокл образ-т зел налеты на коре деревьев. Род Spirogyra - нитчатые водоросли, образ-ие длинные волокна тины в ручьях, речках. Cladophora-в виде ветвящихся "кустиков", прикрепляющихся к камням у берегов рек. Состоящая из многих клеток водяная сеточка (Hydrodictyon), в стоячих водах ("авоська"). У некоторых видов кл-ки соединены в нитчатые колонии. У свободноплавающей колониальной водоросли Scenedesmus серповидные или продолговатые кл-ки объединены в короткие цепочки, род обычен в аквариумах. Самая крупная зел водоросль - морской салат (Ulva), макрофит листовидной формы. Красные водоросли (багрянки): Большинство - морские листовидные, кустистые или корковые макрофиты, обитающие ниже линии отлива. Цвет красный (фикоэритрин), но м б пурпурным или синеватым. Некоторые встречаются в пресной воде. Lemanea - щетковидная форма, в речках и водопадах, талломы прикрепляются к камням. Багрянки не образуют подвижных клеток. Их половой процесс очень сложен, и один жизненный цикл включает несколько фаз. Бурые водоросли: почти все-обитатели моря. Немногие-микроскопич, а среди макрофитов встречаются самые крупные водоросли в мире (ламинарии, фукусы). Все бурые-многоклеточные. Цвет от зел-желтого до темно-коричневого (фукоксантин). Половое размножение-образ-ие подвижных гамет с двумя боковыми жгутиками. Экземпляры, образующие гаметы, часто совершенно не похожи на организмы того же вида, размножающиеся только спорами. Диатомовые водоросли (диатомеи): обширная группа одноклет морских и пресноводных видов.Цвет: желтый, бурый (фукоксантин). Протопласт защищен коробчатой кремнеземной (стеклянной) оболочкой - панцирем (из двух створок, поверхност покрыта хар-ным для вида узором из штрихов, створки пронизаны порами или имеют щель-шов). В клетке находится ядро. Размножение: деление кл-ок надвое и половое. Свободноплав, некоторые прикреплены. Иногда кл-ки объединяются в нити, цепочки или колонии. Различают два типа диатомей: перистые с удлиненными двусторонне-симметричными клетками (они наиболее обильны в пресных водах) и центрические (их больше всего в морях). Панцири после гибели обр-т пористую горную породу - диатомит. Жгутиковые: одноклеточные и подвижные. Кл-ки - зеленые, красные или бесцветные. Нек виды способны к фотосинтезу, др- (сапрофиты) поглощ-ют органику. Половое у некоторых видов. Обычный обитатель прудов - Euglena, зел водоросль с красным "глазком", плавает с помощью жгутика, способна как к фотосинтезу, так и к питанию готовой органикой. В конце лета может окрашивать прудовую воду в красный цвет. Золотистые водоросли: цвет желто-бурый, а кл-ки м б подвиж (жгут) или неподвиж. Размножение бесполое с образ-ем пропитанных кремнеземом цист. Желто-зеленые водоросли: по форме сходны с зел, но отличаются преобладанием специфич желтых пигментов. Их кл стенки состоят из 2 входящих одна в др половинок (у нитчатых видов эти створки в продольном сечении Н-образные). Половое размножение известно лишь у немногих форм. Харовые (лучицы): многоклет, окраска от серовато-зеленой до серой. Кл стенки инкрустированы СаСО3 (отмершие остатки-отложения мергеля). Харофитная структура, 2,5-10 см. Размножение половое. Харовые не близки к какой-либо из перечисленных выше групп,но некоторые считают, что они произошли от зел.
28. Отдел: Цветковые, 240 000 видов, что намного больше всех остальных групп высших растений. Заселяют все климатические зоны земного шара. Их вегетативные и генеративные формы очень изменчивы и способны адаптироваться к разнообразным условиям внешней среды.
Особ-и строения:1.Наличие цветка- репродуктивный орган покрыт-х растений, совмест-й в себе половое и бесполое поколение.2.Дальнейшая редукция мужского и женского гаметофита в жизненном цикле по сравнению с голосеменными. Особенно сильно подвергся изменен женский гаметофит, образ-сь в результате 3-х делений и состоит из 8 клеток (зародышевый мешок), мужской гаметофит состоит из 2-х клеток( пылинка).3.Наличие замкнутой завязи, внутри которой нах-ся семязачатки (замкнутая завязь обесп-ет защиту семязачатков от неблагоприятных условий ).4.Наличие 2-го оплодотворения и образование 3-плоидного эндосперма. У голосеменных эндосперм гаплоидный и образ-ся до оплодотворения. 3-плоидность обеспечивает быстрое развитие эндосперма. 5.Наличие плода, который заключает в себе семена.
Цикл воспроиз-я: покрытосеменные обладают циклом с гетероморфным чредованием поколений и преобладанием спорофита. Мужские гаметофиты переносятся к женским по воздуху (опыление). После попадания пылинки на рыльце пестика генеративная клетка образует 2 спермия, а сифоногенная клетка образует пыльцевую трубку. Пыльцевая трубка проникает в завязь семязачаток внутрь зародыш мешка. Один из спермиев сливается с яйцеклеткой, 2-ой со 2-ым ядром зародыш мешка. Из зиготы зародыш, из 2-го ядра 3-плоидный эндосперм.
Экология: Покрытосеменные являются обширнейшим отделом растительного мира. Более половины всех видов Земных растений относится к этому отделу. Массовое появление покрытосеменных растений отмечается в середине мелового периода (около 120 млн л н). С кайнозойской эры они занимают господствующее положение во флоре Земли, победив в борьбе за существование голосеменные растения. Сильно варьир-т по размерам. Самое маленькое растение-«Вольфия» сем.Рясковые, самое больш- «Эвкалипт». Эфимеры живут несколько недель «веснянка весенняя», а деревья по 1000 лет.
Классификация: В основе подразделения цветковых растений на классы, подклассы, порядки и семейства лежат морфология цветка и соцветия, тонкая структура пыльцевых зёрен, строение и развитие семязачатка, мужск и женск гаметофитов, строение и степень развития зародыша, число семядолей, наличие или отсутствие эндосперма и т.д.Отдел цветковые растения подразделяется на 2 класса: однодольные и 2-ву дольные.
Двудольные - 360 семейств, около 170 тыс. видов.
Подкласс 1. Магнолииды Большей частью древесные растения. Цветки преимущественно обоеполые. Семейства: Магнолиевые (тюльпанное дерево), перечные(мята перечная).
Подкласс 2. Ранункулиды -. Главным образом травы.
Лютиковые(адонис весенний, лютик едкий), Маковые(хохлатка Маршалла).
Подкласс 3. Гамамелидиды образом древесные растения.
Крапивные(крапива двудомная), буковые(дуб черешчатый). Подкласс 4. Кариофиллиды -Обычно травянистые растения, полукустарники или низкие кустарники, редко небольшие деревья. Гвоздичные(звездчатка средняя),гречишные(щавель кислый)Подкласс 5. Дилленииды -Деревья, кустарники или травы. Пионовые(пион узколистный), фиалковые(фиалка трёхцветная) Подкласс 6. Розиды Деревья, кустарники или травы. Розовые(яблоня домашняя), бобовые(клевер красный).
Подкласс 7. Астериды –губоцветные(будра плющевидная), сложноцветные (подсолнечник однолетний)и т.д.
Класс II. Однодольные- Около 70 семейств, свыше 65 тыс. видов.
Подкласс 1. Алисматиды -Водные или болотные травы. Частуховые(подорожник большой), водокрасовые(Элодея).
Подкласс 2. Лилииды. Травы или вторичные древовидные формы. Лилейные(лилия даурская), орхидные(ятрышник).
Подкласс 3. Коммелиниды Травы, иногда с одревесневшим стеблем (бамбуки). Осоковые(камыш озёрный), злаки(кавыль перистый).
Подкласс 4. Арециды Травы или вторичные древовидные формы. Рогозовые
29. Ткани - устойчивые, закон- но повторяющ-ся комплексы клеток, сходные по строению, происхождению и выполняемым функциям. Растительные ткани возникли в процессе эволюции с переходом растений к наземному образу жизни и наибольшей специализации достигли у цветковых. Формирование тканей происходило параллельно с дифференцировкой тела растения на органы. Классификация растительных тканей основана на единстве выполняемых функций, происхождении, сходстве строения и расположении клеток в органах растения.
По строен:1.простые-состоящ из одного типа клеток (апикальн мерист),2.сложные-состоящ из разных типов клеток (эпидерма,проводящ тк).
По происх-ю:1.первичные-образовавш-ся из первичных меристем (прокамбий),2.вторичные-возникшие из вторичных меристем (камбий).
По выполн-м функциям: : 1.образовательные-образ-е тканей корня, стебля, листьев, рост корня и стебля в толщину.
2.покровные-защита органов от высыхания,колебаний температур,повреждений (эпидер)
3.проводящие-провед-е воды и минер вещ-ств из почвы (ксилема, флоэма).
4.ассимиляционные-фотосинтез, газообмен (хлоренхима).
5.механические-сохранение формы растений в условиях воздушной среды (склеренхима).
6.выделительные-выделение из растений продуктов метаболизма (внешние-трихомы, внут секреции млечники, вместилища).
7.запасающие (паренхима)
8.пропускные (эндодерма)
9.всасывающие (эпиблема)
По расположению в теле: 1. Апикальные(верхушечные)-на конце побега или кончике корня. 2. Латеральные-боковые. 3. Интеркалярные-вставочные
5.Механические ткани обеспечивают прочность органов растений. Они составляют каркас, поддерживающий все органы растений, противодействуя их излому, сжатию, разрыву. Основными характеристиками строения механических тканей, обеспечивающими их прочность и упругость, являются мощное утолщение и одревеснение их оболочек, тесное смыкание между клетками, отсутствие перфораций в клеточных стенках.
Механические ткани наиболее развиты в стебле, где они представлены лубяными и древесинными волокнами. В корнях механическая ткань сосредоточена в центре органа.
В зависимости от формы клеток, их строения, физиологического состояния и способа утолщения клеточных оболочек различают два вида механической ткани: колленхиму и склеренхим.
Колленхима представлена живыми паренхимными клетками с неравномерно утолщенными оболочками, делающими их особенно хорошо приспособленными для укрепления молодых растущих органов. Будучи первичными, клетки колленхимы легко растягиваются и практически не мешают удлинению той части растения, в которой находятся. Обычно колленхима располагается отдельными тяжами или непрерывным цилиндром под эпидермой молодого стебля и черешков листьев, а также окаймляет жилки в листьях двудольных. Иногда колленхима содержит хлоропласты.
Склеренхима состоит из вытянутых клеток с равномерно утолщенными, часто одревесневшими оболочками, содержимое которых отмирает на ранних стадиях. Оболочки склеренхимных клеток обладают высокой прочностью, близкой к прочности стали. Эта ткань широко представлена в вегетативных органах наземных растений и составляет их осевую опору.
Различают два типа склеренхимных клеток: волокна и склереиды. Волокна — это длинные тонкие клетки, обычно собранные в тяжи или пучки (например, лубяные или древесинные волокна). Склереиды — это округлые мертвые клетки с очень толстыми одревесневшими оболочками. Ими образованы семенная кожура, скорлупа орехов, косточки вишни, сливы, абрикоса; они придают мякоти груш характерный крупчатый характер.
4.Клетки ассимиляционной ткани содержат хлоропласты и выполняют функцию фотосинтеза. Основная масса этой ткани сосредоточена в листьях, меньшая часть — в молодых зеленых стеблях. В разных органах размеры и форма клеток хлоренхимы неодинаковы. Наиболее разнообразна хлоренхима листьев - мезофилл. Хлоренхима, клетки которой имеют продолговатую форму, называется столбчатой, или палисадной, а состоящая из округлых клеток с большими межклетниками - губчатой. Фотосинтетическая активность хлоренхимы листа пропорциональна числу находящихся в её клетках хлоропластов.Важное значение в структуре хлоренхимы играют межклетники - воздухоносные полости, - резко увеличивающие зелёный экран листа, т.е. площадь соприкосновения клеток мезофилла с воздушной средой. Из воздухоносных полостей хлоренхимы в клетки поступает диоксид углерода, необходимый для фотосинтеза, а в них выделяется кислород.
1.Образовательные ткани, или меристемы, являются эмбриональными тканями. Благодаря долго сохраняющейся способности к делению (некоторые клетки делятся в течение всей жизни) меристемы участвуют в образовании всех постоянных тканей и тем самым формируют растение, а также определяют его длительный рост.
Клетки образовательной ткани тонкостенные, многогранные, плотно сомкнутые, с густой цитоплазмой, с крупным ядром и очень мелкими вакуолями. Они способны делиться в разных направлениях.
По происхождению меристемы бывают первичные и вторичные. Первичная меристема составляет зародыш семени, а у взрослого растения сохраняется на кончике корней и верхушках побегов, что делает возможным их нарастание в длину. Дальнейшее разрастание корня и стебля по диаметру (вторичный рост) обеспечивается вторичными меристемами — камбием и феллоге-ном. Все образовательные ткани делятся митозом.
Образовательные ткани в теле растений располагаются в разных местах, поэтому их делят на следующие группы .1. Верхушечные.2. Латеральные меристемы характерны( для осевых органов).. 3. Интеркалярные, или вставочные, меристемы происходят от верхушечных меристем. Это группы клеток, еще не способных размножаться, однако вставшие на путь дифференциации. Инициальных клеток среди них нет, но много специализированных. 4. Раневые .
6.Выделительные ткани растений - структурные образования, способные выделять из растения или изолировать в его тканях капельно-жидкую воду и продукты метаболизма (секреты). Выделительные ткани растений подразделяются на ткани внутренней и наружной секреции. К первому типу относят разнообразные железистые волоски(представляют собой трихомы, т.е. производные эпидермы, образованные без участия ниже лежащих тканей. Железистые волоски могут быть сидячими, иметь многоклеточную головку и т.д. )и железки(называются эмергенцами, если в их формировании участвует не только эпидерма, но и более глубоко лежащие тканиСолевые железки образуются на листьях, стеблях многих растений солончаков – галофитовОни служат для выведения (против градиента концентрации) на поверхность растения избытка минеральных солей, поступающих через ксилему из почвы, что позволяет растениям переносить очень сильное засоление субстратов)., нектарники(представляют собой разнообразные железистые образования для выделений сахаристой жидкости (нектара) и гидатоды(или водяные устьица - это комплекс клеток в листьях, обеспечивающих выделение из растений капельно-жидкой воды и солей).. Второй тип выделительных тканей включает в себя смоляные каналы (смоляные ходы)- представляют собой длинные трубчатые межклетники, заполненные смолой и окруженные живыми клетками эпителия., вместилища выделений- очень разнообразны по форме, величине и происхождению. Различают схизогенные и лизигенные вместилища. Первые возникают в виде межклетников, заполненных выделенными веществами и окруженных живыми клетками эпителия. Вторые возникают на месте группы клеток, которые распадаются после накопления веществ. Каналообразные выделительные устройства, называются по их содержимому масляными, смоляными, слизевыми и камедевыми ходами., идиобласты- накапливают различные вещества: кристаллы оксалата кальция (одиночные кристаллы, друзы, рафиды и т.д.), слизи, таннины, эфирные масла. Они встречаются среди клеток разных тканей, могут иметь разнообразную форму и химический состав. (специализированные клетки), млечники (млечные трубки)-млечные трубки) выполняют разнообразные функции - проводящую, запасающую, выделительную. Стенка их состоит из целлюлозы. Это живые клетки с цитоплазмой, многими ядрами и вакуолью, заполненной млечным соком (латексом- млечный сок каучуконосных растений. Различают два вида млечников: членистые и нечленистые. Членистые образуются так же, как сосуды, в результате разрушения поперечных стенок у вертикального ряда клеток. Нечленистые млечники возникают в результате разрастания специальных клеток зародыша. Млечники располагаются или только во флоэме, или пронизывают весь орган (стебель, корень, лист).
3.Проводящие ткани обеспечивают передвижение воды и растворенных в ней питательных веществ по растению. Различают два вида проводящей ткани — ксилему (древесину) и флоэму (луб).
Ксилема —это главная водопроводящая ткань высших сосудистых растений, обеспечивающая передвижение воды с растворенными в ней минеральными веществами от корней к листьям и другим частям растения (восходящий ток). Она также выполняет опорную функцию. В состав ксилемы входят трахеиды и трахеи (сосуды) , древесинная паренхима и механическая ткань.
Трахеиды представляют собой узкие, сильно вытянутые в длину мертвые клетки с заостренными концами и одревесневшими оболочками. Проникновение растворов из одной трахеиды в другую происходит путем фильтрации через поры — углубления, затянутые мембраной. Жидкость по трахеидам протекает медленно, так как поровая мембрана препятствует движению воды. Трахеиды встречаются у всех высших растений, а у большинства хвощей, плаунов, папоротников и голосеменных служат единственным проводящим элементом ксилемы. У покрытосеменных растений наряду с трахеидами имеются сосуды.
Трахеи (сосуды) —это полые трубки, состоящие из отдельных члеников, расположенных друг над другом. В члениках на поперечных стенках образуются сквозные отверстия — перфорации, или эти стенки полностью разрушаются, благодаря чему скорость тока растворов по сосудам многократно увеличивается. Оболочки сосудов пропитываются лигнином и придают стеблю дополнительную прочность. В зависимости от характера утолщения оболочек различают трахеи кольчатые, спиральные, лестничные и др.
Флоэма проводит органические вещества, синтезированные в листьях, ко всем органам растения (нисходящий ток). Как и ксилема, она является сложной тканью и состоит из ситовидных трубок с клетками-спутницами , паренхимы и механической ткани. Ситовидные трубки образованы живыми клетками, расположенными одна над другой. Их поперечные стенки пронизаны мелкими отверстиями, образующими как бы сито. Клетки ситовидных трубок лишены ядер, но содержат в центральной части цитоплазму, тяжи которой через сквозные отверстия в поперечных перегородках проходят в соседние клетки. Ситовидные трубки, как и сосуды, тянутся по всей длине растения. Клетки-спутницы соединены с члениками ситовидных трубок многочисленными плазмодесмами и, по-видимому, выполняют часть функций, утраченных ситовидными трубками (синтез ферментов, образование АТФ).
Ксилема и флоэма находятся в тесном взаимодействии друг с другом и образуют в органах растения особые комплексные группы — проводящие пучки.
Покровные ткани располагаются на поверхности всех органов растения. Они выполняют главным образом защитную функцию — защищают растения от механических повреждений, проникновения микроорганизмов, резких колебаний температуры, излишнего испарения и т. п. В зависимости от происхождения различают три группы покровных тканей —эпидермис, перидерму и корку.
Эпидермис (эпидерма, кожица) — первичная покровная ткань, расположенная на поверхности листьев и молодых зеленых побегов . Она состоит из одного слоя живых, плотно сомкнутых клеток, не имеющих хлоропластов. Наружная поверхность клеток этой ткани часто одета кутикулой или восковым налетом, что является дополнительным защитным приспособлением. В эпидерме листьев и зеленых стеблей имеются устьица, которые регулируют транспирацию и газообмен растения. (трихомы и железистые волоски).
Перидерма — вторичная покровная ткань стеблей и корней, сменяющая эпидермис у многолетних (реже однолетних) растений . Ее образование связано с деятельностью вторичной меристемы —феллогена (пробкового камбия), клетки которого делятся и дифференцируются в центробежном направлении (наружу) в пробку (феллему), а в центростремительном, (внутрь) — в слой живых паренхимных клеток (феллодерму). Пробка, феллоген и феллодерма составляют перидерму.
Клетки пробки пропитаны жироподобным веществом — суберином —и не пропускают воду и воздух, поэтому содержимое клетки отмирает и она заполняется воздухом. Многослойная пробка образует своеобразный чехол стебля, надежно предохраняющий растение от неблагоприятных воздействий окружающей среды. Для газообмена и транспирации живых тканей, лежащих под пробкой, в последней имеются особые образования —чечевички; это разрывы в пробке, заполненные рыхло расположенными клетками.
Корка образуется у деревьев и кустарников на смену пробке. В более глубоко лежащих тканях коры закладываются новые участки феллогена, формирующие новые слои пробки. Вследствие этого наружные ткани изолируются от центральной части стебля, деформируются и отмирают, На поверхности стебля постепенно образуется комплекс мертвых тканей, состоящий из нескольких слоев пробки и отмерших участков коры. Толстая корка служит более надежной защитой для растения, чем пробка.
Запасающие ткани несут функции накопления и хранения запасов воды и органических веществ. Запасы хранятся в течение длительного срока (как, например, зимние запасы крахмала в клубнях и корневищах) либо потребляются и вновь пополняются в период вегетации (как, например, запасы воды в листьях и стеблях мясистосочных растений).
Ткани, запасающие воду (водоносные ткани), состоят либо из живых паренхимных клеток с тонкими целлюлозными иногда ослизняющимися стенками, либо из мертвых трахеидоподобных клеток, отличающихся от обычных трахеид крупными размерами и почти изодиаметрической формой. Водоносную ткань первого рода можно видеть, например, в листьях эпифитной пеперомии (Peperomia incana). Она состоит здесь из нескольких слоев живых тонкостенных клеток с крупной центральной вакуолей, заполненной водянистым клеточным соком. При подсыхании листа вода из клеток водоносной ткани передается другим тканям, главным образом ассимиляционным. Стенки водоносных клеток сокращаются, при этом сначала равномерно, а при дальнейшей отдаче воды в радиальных стенках образуются складки; когда лист снова обогащается водой, клетки принимают прежнюю форму. Мощная внутренняя водоносная ткань имеется у суккулентов (растений с сочными, богатыми водой вегетативными органами): у одних - преимущественно в листьях (у агав, алоэ, молодил Sempervivum), у других - в стеблях (у кактусов, некоторых молочаев Euphorbia).
Среди тканей, запасающих пластические вещества, различают два типа: 1) ткани, накопляющие запасы в полостях клеток, и 2) ткани с запасами в полостях клеток и в оболочках их. В тканях первого типа оболочки обычно тонки и снабжены мелкими простыми порами. Клетки содержат запасные вещества (чаще всего - сахара, крахмал, инулин, аминокислоты, белки): а) в растворе (в корнеплоде свеклы, в сочных чешуях луковицы лука); б) частью в растворе (амиды, белковые вещества), частью в твердом виде (крахмал, кристаллиды белка - в клубнях картофеля); в) в твердом (алейроновые зерна) и полужидком (капли жирных масел) виде (в эндосперме клещевины); г) почти полностью в твердом виде (в богатых алейроновыми зернами и крахмалом семядолях фасоли, гороха, чечевицы). Запасающие ткани второго рода содержат обычно в клеточных полостях алейроновые зерна и жирные масла, а в оболочках клеток - отложения гемицеллюлоз либо амилоидов, а иногда слизей. Оболочки клеток сильно утолщаются и снабжаются многочисленными крупными простыми порами. Замыкающие пленки пор, а иногда и утолщенные участки клеточных стенок пронизываются плазмодесменными канальцами. Запасающие ткани этого рода содержатся в эндосперме семян кофе, финиковой пальмы, в семядолях люпинов (Lupinus), недотрог (Impatiens).
Расходование растением запасных веществ подготовляется гидролизом их (при участии ферментов), приводящим к образованию растворимых в воде веществ, перемещающихся в места использования - в распускающиеся почки, в развивающийся проросток, в созревающие плоды и семена. Запасные вещества расходуются растением на рост, развитие, воспроизведение1 и на дыхание, освобождающее рабочую энергию.
9.Эпибле́ма, или ризоде́рма — первичная покровная ткань молодых корней растений. Формирует корневые волоски.
Клетки эпиблемы возникают из самого наружного слоя корневой меристемы — дерматогена. Эпиблема покрывает корень на протяжении от корневого чехлика до зоны проведения, длина её варьирует в пределах от нескольких миллиметров до нескольких сантиметров. Наибольшего развития клетки эпиблемы достигают в зоне поглощения корня, где формируют бугорки, вскоре значительно выпячивающиеся наружу — корневые волоски. Их количество довольно велико, они покрывают всю эпиблему на протяжении зоны проведения и обеспечивают растение большей частью воды и минеральных солей, получаемых из окружающей среды.
По мере роста корня эпиблема постепенно разрушается. Её клетки слущиваются, обнажая опробковевшую экзодерму, выполняющую функцию защитной ткани. В отличие от пробки, клетки экзодермы могут сохранять проницаемость оболочек, а из части клеток экзодермы возможно формирование пропускных клеток, позволяющих передвижение веществ.
Эндодерма (от эндо... и греч. dérma — кожа, оболочка), внутренний однорядный слой плотно сомкнутых паренхимных клеток первичной коры, прилегающий к центральному цилиндру осевых органов высших растений. В стеблях Эндодерма мало дифференцирована, обычно содержит вторичный крахмал. В корнях радиальные и поперечные стенки клеток Эндодерма имеют утолщения в виде поясков, содержащие суберин и лигнин, тонкостенными остаются пропускные клетки этого слоя. Т. о., Эндодерма является физиологическим барьером, регулирующим поступление воды и ионов из первичной коры в центральный цилиндр корня.
Отлич-е растит ткан от животн-х:
1.Для клеток растений х-но обратимость процесса дифференцировки. Большинство клеток раст-й, сохран-х живой протопласт при определн-х условиях вновь могут приобрести способность к делению и превращению в другие типы клеток (из любой клетки можно получить целое растение).
2.У растит клетки нет генетич-и запрограм-го механизма, огранич-щего число деления.
Апопласт-совокупность межклетн-в, клеточных оболочек и полостей отмерших клеток (неживая состав-щая растений).Значение:механическая функция,проведение воды,защита.(Перидерма) Транспорт вещ-ств происходит без активного участия самой клетки.Симпласт-совокупность протопластов и связывающих их плазмодесм (живая составляющая тела растений). Транспорт по симпласту происходит быстрее, чем диффузия (1-6 см/ч), и не требует затраты энергии
30. Протисты или простейшие— гетерогенная группа эукариотических живых организмов.(~ 30 000 видов)
Простейшие, как правило, микроскопических размеров, обитают в воде, влажной почве или в теле различных животных и человека. Форма их тела весьма разнообразна— от неопределённой (как у амёбы) до удлинённой, обтекаемой, веретеновидной (трипаносома), некоторые имеют наружную раковину (фораминиферы).
Тело большинства простейших состоит из одной клетки, содержащей одно или несколько ядер. Клетка простейших типична для эукарионых организмов и состоит из цитоплазмы и одного или нескольких ядер. Цитоплазма ограничена снаружи трехслойной мембраной. В цитоплазме простейших различают наружный, более прозрачный и плотный слой— эктоплазму и внутренний, зернистый слой— эндоплазму. В эндоплазме сосредоточены все основные органеллы клетки: ядро, митохондрии, рибосомы, лизосомы, эндоплазматическая сеть, аппарат Гольджи и др. Кроме того, у простейших имеются особые органеллы: опорные, сократительные фибриллы, пищеварительные и сократительные вакуоли и др. Ядро покрыто двухслойной мембраной с порами. Внутри ядра находится кариоплазма, в которой распределены хроматин и ядрышки. Хроматин представляет собой деспирализованные хромосомы, состоящие из ДНК и белков типа гистонов. Ядрышки подобны рибосомам и состоят из РНК и белков. Ядра простейших разнообразны по составу, форме, размерам.
Простейшие способны передвигаться с помощью ложноножек, жгутиков или ресничек, реагируют на различные раздражения (фототаксис, хемотаксис, термотаксис.). Питаются простейшие мельчайшими животными, растительными организмами и гниющими органическими веществами. Пути поступления пищи в организм клетки также различны: пиноцитоз, фагоцитоз, осмотический путь, активный перенос веществ через мембрану. Поступившую пищу они переваривают в пищеварительных вакуолях, заполненных пищеварительными ферментами. Некоторые из них, имеющие фотосинтезирующих внутриклеточных симбионтов— хлорелл или хлоропласты (эвглены) способны синтезировать органическое вещество из неорганических веществ с помощью фотосинтеза.
Газообмен у простейших осуществляется всей поверхностью тела осмотическим путем; выделение продуктов обмена веществ и избытка воды происходит через поверхность тела, а также с помощью специальных периодически образующихся сократительных (или пульсирующих) вакуолей.
Размножение простейших происходит бесполым и половым путём в зависимости от условий существования. При бесполом размножении сначала ядро делится на две или несколько частей, а затем делится цитоплазма на две (равные или неравные) или много частей (соответственно числу вновь образовавшихся ядер). В результате из одного организма образуется два (одинаковых или неравных по величине) или несколько новых организмов. При половом размножении две равные или различные по величине и строению (мужская и женская) особи сливаются друг с другом, образуя зиготу, которая затем начинает размножаться бесполым путём. Иногда между двумя особями происходит обмен частью ядер при соприкосновении особей (образования зиготы не наблюдается).
При неблагоприятных условиях простейшие способны образовывать цисты.
Классиф-ия:
Тип:
1.Саркомастигофоры (амёбы, лучевики),2.Апикомплексы (малярийный плазмодий),3.Микроспоридии,4.Миксоспоридии,5.Инфузории,6.Лабиринтулы,7.Ацетоспоридии.
31. Млекопитающие — класс позвоночных животных, основными отличительными особенностями которых являются живорождение (за исключением клоачных) и вскармливание детёнышей молоком (насч. ~ 5000 видов). В ходе эволюции млекопитающие осуществили широчайшую адаптивную радиацию, т.е. приспособились к самым разным экологическим нишам.
Характерные особенности:
1. Наличие волосяного покрова (шерсти), потовых и сальных желёз,2 . особый тип строения головного мозга (в том числе сильное развитие конечного мозга, переход к нему функций основного зрительного центра и центра управления сложными формами поведения),3. наличие трёх слуховых косточек среднего уха, наружного ушного прохода и ушной раковины,4. семь позвонков в шейном отделе позвоночника,5. теплокровность,6. 4-х камерное сердце,7. альвеолярное строение лёгких, 8. зубы сидящие в ячейках (альвеолах) челюстей; гетеродонтность (разнозубость),9. безъядерные эритроциты.
Особенности строения: Подавляющее большинство млекопитающих одето волосяным покровом, который служит прежде всего для сохранения внутреннего тепла животного. Отсутствие волосяного покрова у некоторых млекопитающих всегда является следствием его редукции под влиянием определенных условий среды. В коже млекопитающих заложены различные кожные железы: наиболее обычны из них потовые и сальные железы, играющие важную роль в терморегуляции. Видоизмененными потовыми железами являются молочные железы.
Скелет взрослых млекопитающих состоит в основном из костных элементов. Позвоночник образован платицельными позвонками, имеющими плоские сочленовные поверхности, между которыми расположены упругие мениски. Позвоночник делится на шейный, грудной, поясничный, крестцовый и хвостовой отделы. Шейных позвонков у млекопитающих всегда семь. Хорошо развиты ребра; вместе с грудными позвонками и грудиной они образуют грудную клетку. Крестцовые позвонки сливаются в единую кость — крестец. Конечности пятипалого типа, но у многих видов некоторые пальцы редуцированы. Скелет передних конечностей состоит из плечевой кости, двух костей предплечья (локтевой и лучевой), ряда мелких косточек запястья, вытянутых косточек пясти и фаланг пальцев. Плечевой пояс образован лишь лопатками и ключицами: коракоидная кость в онтогенезе прирастает к лопатке, образуя ее коракоидный отросток.
Скелет задних конечностей включает бедренную кость, две кости голени (большую и малую берцовые), ряд костей предплюсны, кости плюсны и фаланг пальцев. Тазовый пояс состоит из трех пар костей: седалищных, лобковых и подвздошных; у взрослых особей большинства видов они слиты в единую безымянную кость.
Череп млекопитающих сочленяется с первым позвонком — атлантом — двумя затылочными мыщелками. Для черепа млекопитающих характерна тенденция слияния в процессе онтогенеза отдельных костей друг с другом. Отмечается большая величина мозговой коробки, что связано с крупными размерами мозга. Нижняя челюсть простого строения — она образована одной зубной костью. Квадратная и сочленовная кости черепа пресмыкающихся превращаются у млекопитающих в слуховые косточки (наковальню и молоточек). Для головного мозга млекопитающих характерно сильное развитие коры больших полушарий, образованной серым мозговым веществом. Большой мозжечок состоит из двух полушарий, соединенных поперечной комиссурой. От головного мозга отходит 12 пар головных нервов. Пищеварительный тракт млекопитающих распадается на ряд хорошо обособленных отделов: ротовую полость, глотку, пищевод, простой или сложный желудки, тонкие и толстые кишки. Имеется ряд пищеварительных желез — печень, поджелудочная и другие. Клоака свойственна только низшим представителям этого класса (яйцекладущим).
Зубы зверей сидят в альвеолах верхней и нижней челюстей. Каждый из них состоит из выступающей наружу коронки и находящегося в альвеоле корня; часто между коронкой и корнем имеется перехват — шейка зуба. Коронка образована дентином, покрытым сверху слоем очень твердой эмали. Корень зуба также состоит из дентина, но снаружи одет зубным цементом. Внутри зубов обычно расположена полость, наполненная мягкой пульпой.
Органами дыхания млекопитающих служат легкие альвеолярного строения. Дыханию способствуют движения диафрагмы, разделяющей грудную и брюшную полости.
Имеется два круга кровообращения. Сердце четырехкамерное, что предупреждает смешение артериальной и венозной крови. Из двух дуг аорты эмбрионов у взрослых особей сохраняется только левая.
Почки вторичные, тазовые, обычно бобовидной формы. Все млекопитающие раздельнополые животные. У многих из них хорошо выражен половой диморфизм. Почти все млекопитающие родят живых детенышей; лишь утконосы, ехидны размножают откладывая яйца. Классификация
Воспроизведение: Млекопитающие — раздельнополые животные. В органах размножения самки — яичниках — развиваются яйцеклетки, в органах размножения самца — семенниках — сперматозоиды. Оплодотворение у млекопитающих внутреннее. Созревшие клетки поступают в парный яйцевод, где происходит их оплодотворение. Оба яйцевода открываются в особый орган женской половой системы — матку, которая есть только у млекопитающих. Матка представляет собой мускулистый мешок, стенки которого способны сильно растягиваться. Начавшаяся делиться яйцеклетка прикрепляется к стенке матки, и все дальнейшее развитие плода происходит в этом органе. В матке оболочка зародыша плотно соприкасается с ее стенкой. В месте соприкосновения образуется детское место, или плацента. Зародыш соединен с плацентой пуповиной, внутри которой проходят его кровеносные сосуды. В плаценте через стенки сосудов из крови матери в кровь зародыша поступают питательные вещества и кислород и удаляются углекислый газ и другие вредные для зародыша продукты жизнедеятельности. Длительность развития зародыша в матке у разных млекопитающих различна (от нескольких дней до 1,5 лет). На определенной стадии зародыш млекопитающих имеет зачатки жабр и по многим другим признакам сходен с зародышами земноводных и пресмыкающихся.
Классификация: Существует много различных классификаций млекопитающих. Ниже приведён только один из возможных вариантов, в котором класс млекопитающих делится на два подкласса, шесть инфраклассов (три из которых вымерли) и множество отрядов (слово «отряд» не указывается).
Подкласс первозвери
Инфракласс клоачные
Однопроходные: ехидны и утконосы
Подкласс звери,
Инфракласс сумчатые,
опоссумы,
малобугорчатые,
микробиотерииды,
кроты сумчатые,
хищные сумчатые
бандикутообразные,
двурезцовые,
кенгуру, коала, вомбаты.
Инфракласс плацентарные или высшие звери,
Неполнозубые,: муравьеды, ленивцы, броненосцы и др.
Панголины или ящеры, 7 современных видов
Насекомоядные, ежи, кроты, землеройки и др.
Прыгунчики14-15 видов
Тенрекообразные,: тенреки и златокротовые
Грызуны, мыши, крысы, белки, бобры, хомяки и многие другие
Зайцеобразные, зайцы, кролики, пищухи
Трубкозубые, единственный современный вид — африканский трубкозуб
Шерстокрылы или кагуаны, только два современных вида
Рукокрылые,: летучие мыши и крыланы
Тупайеобразные,: тупайи
Приматы, лемуры и обезьяны (в том числе человек)
Хищные,: кошки, собаки, медведи, куницы, еноты и др.
Ластоногие,: моржи, тюлени, морские слоны и др.
Китообразные, дельфины, киты, кашалоты
Непарнокопытные или непарнопалые, лошади, зебры, носороги и др.
Парнокопытные или парнопалые,: олени, коровы, свиньи, антилопы и многие другие
Даманы или жиряки,: 7-12 современных видов
Хоботные, слоны и много вымерших видов, включая мамонтов и мастодонтов
Сирены,: ламантины и дюгони
32. Главная цель охраны природы - поддержание уровня биологического разнообразия, которое обеспечивает эволюцию популяций, а также экосистем разного ранга - от биогеоценозов до биосферы. К природоохранным мероприятиям относят восстановление используемых человеком биоресурсов. Сохранение биологического разнообразия является составной и неотъемлемой частью стратегии устойчивого развития. Разнообразие живых существ используется человеком и для введения новых видов живых существ в культуру и в селекционном процессе.Для сохранения биологического разнообразия необходим представительный набор охраняемых территорий, совершенствование охраны отдельных видов живых существ и экосистем, их всестороннее исследование, а также использование различных методов сохранения видов и генофонда, как в природе, так и в искусственных условиях. Принципы охраны природы. Хозяйственная деятельность вызывает в природе многочисленные изменения, последствия которых необходимо уметь прогнозировать. В процессе длительного использования природных ресурсов были разработаны общие принципы и правила рационального использования и охраны природы. 1.принцип- все явления природы имеют для человека множественное значение и должны оцениваться с разных точек зрения. К каждому явлению необходимо подходить с учетом интересов разных отраслей производства и сохранения восстановительной силы самой природы. Так, лес рассматривается прежде всего как источник древесины и химического сырья, однако леса имеют водорегулирующее, почвозащитное, климатообразующее значение. Лес важен как место отдыха людей. В этих случаях промышленное значение леса отодвигается на второй план. 2.принцип - в необходимости строгого учета местных условий при использовании и охране природного ресурса. Его называют правилом региональности. Особенно это касается использования водных и лесных богатств. На Земле много мест, где сейчас ощущается дефицит пресной воды. Избыток воды в других местах не улучшает затруднительного положения с водой в засушливых районах. Нет ничего более губительного, чем интенсивное расходование ресурса там, где он в недостатке, на основании того, что в других местах этот ресурс находится в избытке. Согласно правилу региональности обращение с одним и тем же природным ресурсом в разных районах должно быть различным и зависеть от того, как этот ресурс в данной местности представлен в настоящее время. 3.принцип- охрана одного объекта означает одновременно охрану и других объектов, тесно с ним связанных. Охрана водоема от загрязнения — это одновременная охрана рыб, обитающих в нем. Часто в природе складываются отношения противоположного характера, когда охрана одного объекта приносит вред другому. Поэтому охрана каждого природного объекта должна быть соотнесена с охраной других. Следовательно, охрана природы должна быть комплексной. Охраняться должна не сумма отдельных природных ресурсов, а природный комплекс (экосистема), включающий различные компоненты, соединенные естественными связями, сложившимися в процессе длительного исторического развития. Правовой основой охраны природы К. О. являются Конституция РФ, международные договоры, законы об охране природы и основных компонентов окружающей природной среды (об особо охраняемых природных территориях К.О.,о порядке использования лесов на территории К.О.), постановления законодательных учреждений разного уровня. Высшую юридическую силу, прямое действие и применение на всей территории России имеет Конституция РФ. В настоящее время действует Закон Российской Федерации «Об охране окружающей природной среды». Основой его является признание природы и ее богатств «национальным достоянием народов России, естественной основой их социально-экономического развития и благосостояния человека».
ООПТ. До июля 2009 года в К.О. было 80 ООПТ. Гос заповедник-1, заказники общего профиля-19, 3 ботанических и 16 зоологических памятников природы, 1 дендропарк, лечебно-оздоровительная местность-1. В связи с тем, что ООПТ регионального значения были образованы с нарушением законодательства (отсутвовало комплексное эко обследование и эко экспертиза) постановлением администрации К.О. в июле 2009 года снят статус с 75 ООПТ К.О. В настоящее время статус ООПТ имеют: 1 лечебно-оздоровительная местность регионального значения пушкаро-жадинское месторождение лечебных торфов в Кореневском районе; 2 памятник природы погребенная микулинская палеобалка в карьере александровского месторождения суглинков в Курском районе; 3 дендрологический парк г. Железногорск. ЦЧГЗ-федерального статуса ООПТ общей площадью более 5 тыс га.
Для обеспечения охраны редких и находящихся под угрозой исчезновения диких животных, дикорастущих растений и грибов, обитающих (произрастающих) на территории Курской области, учредить Красную книгу Курской области.
(Закон Курской области "О Красной книге Курской области" (принят Курской областной Думой 2004))
. Красная книга Курской области является официальным документом, содержащим свод сведений об объектах животного и растительного мира, а также необходимых мерах по их охране и восстановлению.Специально уполномоченным органом по ведению Красной книги Курской области был определен департамент экологической безопасности и природопользования Курской области.В Красную книгу Курской области внесено 119 объектов фауны(сурок степной,выхухоль) и 212 объектов флоры(кавыль перистый, волчеягодник боровой, проломник Козо-Полянского).Ежегодно на территории области проводятся работы по учету, исследованиям, охране и обследованию объектов фауны и флоры.
Издание Красной книги Курской области осуществляется не реже одного раза в 10 лет.