

Гиперфосфатемия
Гиперфосфатемия – состояние, при котором концентрация фосфатов в сыворотке крови превышает 5 мг / 100 мл у взрослых В клинической практике наблюдается достаточно часто. Самой распространенной причиной этого состояния является хроническая почечная недостаточность, обусловливающая сниженную экскрецию фосфатов с мочой. Кроме того, гиперфосфатемия может быть следствием увеличенного поступления фосфатов во внеклеточную жидкость или их повышенного поступления в организм .
Причины гиперфосфатемии
I. Снижение почечной экскреции фосфатов:
почечная недостаточность (острая, хроническая); гипопаратиреоз; псевдогипопаратиреоз; циркуляция аномального ПТГ; акромегалия; опухолевый кальциноз.
II. Повышенное поступление фосфатов во внеклеточную жидкость:
опухолевые заболевания (лейкемия, лимфома); повышенный катаболизм; респираторный ацидоз.
III. Введение солей РО4 или витамина D:
введение метаболитов витамина D в фармакологических целях;
поступление внутрь и (или) введение солей фосфатов.
IV. Смешанные состояния:
кортикальный гиперостоз;
перемежающаяся гиперфосфатемия;
Большая часть клинических эффектов гиперфосфатемии связано со вторичными изменениями метаболизма кальция.
ПАТОЛОГИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Гормоны щитовидной железы (тиреоидные гормоны) представляют собой йодированные молекулы, называемые йодтиронинами; это трийодтиронин (Т3) и тетрайодтиронин (Т4 или тироксин). Синтез, запасание и секреция этих гормонов включают следующие стадии:
1. Накопление фолликулярными клетками щитовидной железы йодида, поступающего в организм с пищей и водой. Процесс обеспечивается работой йодидного насоса на базальной мембране клеток;
2. Окисление йодида в клетках под действием пероксидазной системы, локализованной вблизи апикальной мембраны;
3. «Организация» активного йода в виде присоединения его к молекулам аминокислоты тирозина;
4. Йодирование тирозильных остатков по одному или двум возможным участкам молекулы с образованием моно- и дийодтирозинов (Т1 и Т2), остающихся в составе тиреоглобулина;
5. Изменение структуры тиреоглобулина, обеспечивающее возможность конденсации различных йодированных тирозильных остатков (2+1 или 2+2). Образование в результате этой реакции три- и тетрайодированных тирониновых групп (Т3 и Т4), все еще остающихся в составе тиреоглобулина;
6. Секреция тиреоглобулина, содержащего йодтиронины, в просвет фолликулов и его запасание в них в виде коллоида;
7. Поступление коллоида из просвета фолликул в фолликулярные клетки путем эндо-цитоза при участии внутриклеточной сети микротрубочек;
8. Расщепление тиреоглобулина ферментами лизосом и поступление его йодированных компонентов в цитоплазму;
9. Дейодирование моно- и дийодтирозинов цитоплазматическими дегалогеназами и использование образующихся при этом молекул тирозина и йодида для повторного синтеза тиреоглобулинов;
10. Секреция три- и тетрайодтиронина в кровь.
II. Главным стимулятором секреции Т4 и Т3 является тиреотропный гормон (ТТГ). Под контролем этого гормона находятся процессы, обеспечивающие синтез, запасание и секрецию йодтиронинов, включая: работу йодидного насоса, организацию активного йода, синтез тиреоглобулина, реакцию конденсации, поглощение коллоидных капель путем эндоцитоза, их слияние с лизосомами, распад тиреоглобулина с высвобождением Т3 и Т4 в кровь. Все эти процессы стимулируются ТТГ.
В свою очередь, секреции ТТГ контролируется двумя механизмами (рис. .):
1. Пептидным гормоном гипоталамуса - тиролиберином, стимулирующим синтез и секрецию ТТГ в аденогипофизе;
2. Тиреоидными гормонами, непосредственно ингибирующими секрецию ТТГ по механизму отрицательной обратной связи.;
3. Кроме тиролиберина и тиреоидных гормонов на секрецию ТТГ прямо или косвенно влияют и многие другие факторы, но их роль менее значительна.
III. Транспорт тиреоидных гормонов (Т3 и Т4) в крови, в основном, осуществляется в связанной с белками плазмы форме. Тироксин обладает высоким сродством к гликозилированному глобулину, называемому тиронинсвязывающим глобулином (ТСГ). Из общего количества Т4 с ТСГ связываются 75%. 15-20% тироксина связываются с преальбумином {тирок-синсвязывающий npеальбумин, ТСПА), и менее 0,05% находится в несвязанном состоянии.
IV. Периферические превращения тироксина связаны с его дейодированием и образованием различных соединений, в структуре которых содержится меньше молекул йода. Самым главным из таких превращении является конверсия Т4 более активный T3.
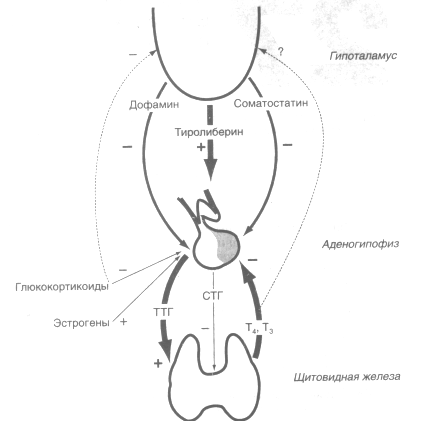
Рис. . Регуляция гипоталамо-гипофизарно-тиреоидной системы
V. Механизм действия тиреоидных гормонов реализуется через их связывание со специфическими внутриклеточными рецепторами (рис. .). Этому способствует их высокая способность к довольно легкому проникновению через плазматические мембраны клеток-мишеней (отчасти с помощью процессов активного транспорта).
Хотя единый механизм действия тиреоидных гормонов пока неизвестен, важнейшую роль в нем играет прямая стимуляция процессов транскрипции в ядре, приводящая к усилению внутриклеточного синтеза белка.
В настоящее время выделяют ядерные и неядерные механизмы действия тиреоидных гормонов. Среди них: прямая стимуляция синтеза белка в митохондриях, приводящая к увеличению их размеров и возрастанию активности ферментов, в том числе - ферментов дыхательной цепи;прямое действие на транспортные механизмы клеточных мембран (например, на транспорт аминокислот) возможно в результате образования цАМФ или стимуляции мембранной АТФ-азы.
I. Mетаболическиe эффекты тиреоидных гормонов проявляются прежде всего в возрастании скорости метаболизма и, соответственно, величины основного обмена Поскольку КПД реакций, протекающих с использованием молекул АТФ, относительно низкий (5-30%), их возросшее потребление приведет к росту теплопотерь, что и будет определять калоригенный эффект тиреоидных гормонов.
Влияние тиреоидных гормонов на белковый обмен проявляется возрастанием скорости как синтеза, так и катаболизма белков, при большей выражениости катаболических процессов.
Результирующей этих сдвигов в условиях повышенной продукции йодтиронинов является развитие отрицательного азотистого баланса и атрофия мышц, сопровождающаяся потерей массы тела .
При пониженной продукции тиреоидных гормонов (гипотиреоз) скорость катаболизма белков уменьшается, что приводит к положительному азотистому балансу. Но снижение катаболизма азотсодержащих веществ сопровождается накоплением в соединительной ткани коллагена и избытком гликозаминогликанов , что способствует формированию измененных гипергидратированных структур. В результате развивается т.н. «слизистый отек » - микседема - признак, характерный для тиреоидной недостаточности. Считается, что через участие и белковом обмене тиреоидные гормоны играют существенную роль в обеспечении нормального роста, развития и целостности тканей.
Влияние тиреоидных гормонов на углеводный обмен реализуется активацией его различных сторон. В частности, йодтиронины стимулируют глюконеогенез, аэробный и анаэробный пути катаболизма глюкозы- поглощение глюкозы мышцами и другими периферическими тканями за счет активации механизмов ее транспорта через плазматическую мембрану . Вместе с тем толерантность к глюкозе в условиях повышенной продукции тиреоидных гормонов снижается, что обусловлено как возросшим всасыванием и образованием самой глюкозы( глюконеогенез), так и редуцирующим влиянием тиреоидных гормонов на экспрессию клеточных рецепторов к инсулину на адипоцитах. В результате гипергликемия и глюкозурия могут присутствовать при гипертиреоидных состояниях. При дефиците тиреоидных гормонов (гипотиреоз) изменения в обмене глюкозы носят противоположную направленность.
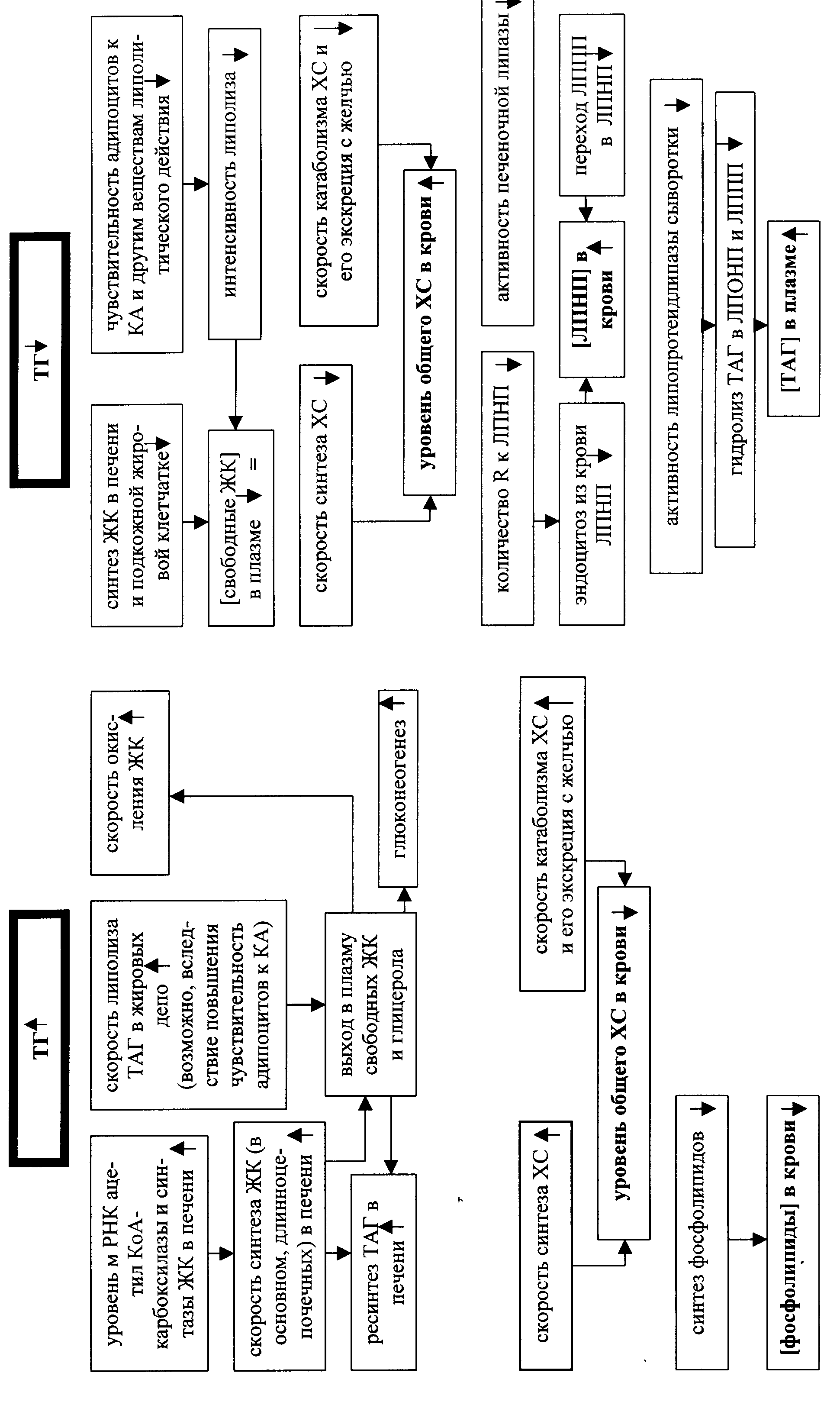
Влияние тиреоидных гормонов на липидный обмен проявляется активацией липолиза, скорости окисления жирных кислот при одновременном повышении их использования для синтеза триацилглицеридов (ТАГ) в печени. Однако, преобладание католического эффекта обусловливает при повышенной продукции йодтиронинов общее уменьшение запасов жира в организме, похудание и снижение концентрации липидов (триацилглицеридов, холестерина, фосфолипидов) в крови (рис. ), При снижении уровня тиреоидных гормонов концентрация липидов в крови возрастает. Причем, это возрастание (например, холестерина) определяется не усилением его синтеза, который на самом деле снижен, а еще более значимым падением скорости катаболизма. Тот же механизм лежит в основе повышения ТАГ в крови при гипотиреоидном состоянии. Это повышение обусловлено снижением активности печеночной липазы и липопротеидлипазы сыворотки, принимающей участие в обороте ТАГ.
Влияние тиреоидных гормонов на минеральный обмен определяется их способностью стимулировать как резорбцию, так и синтез костной ткани, но первый процесс (резорбция) усиливается в значительно большей степени. Таким образом, при избытке Т3 или Т4 наблюдается деминерализация костей с повышением риска переломов, гиперкальциемия, потеря кальция и фосфора с мочой и калом. Кроме того, тиреоидные гормоны ускоряют кругооборот белкового (коллагенового) матрикса кости.
VII. Влияние тиреоидных гормонов на функциональные системы организма наиболее значимо проявляется в отношении центральной нервной системы и системы кровообращения.
Йодтиронины абсолютно необходимы для нормального психического развития. При их недостаточности в период между рождением и половой зрелостью ребенок остается не только физически, но и умственно отсталым {кретинизм) из-за недостаточного развития межклеточных контактов в головном мозге, гипоплазии нейронов коры больших полушарий и задержки миелинизации нервных волокон. Тиреоидные гормоны необходимы для развития нервной системы не только на последних месяцах беременности и в постнатальном периоде, но и на самых ранних этапах нейро-и глиогенеза. В это время щитовидная железа плода еще не способна полностью синтезировать гормоны и обеспечение ими осуществляется за счет гормонов матери, которые в определенных количествах cпособны проходить, через плаценту. Гипотериоз у взрослых характеризуется замедлением умственных процессов, ослаблением памяти, потерей инициативности.
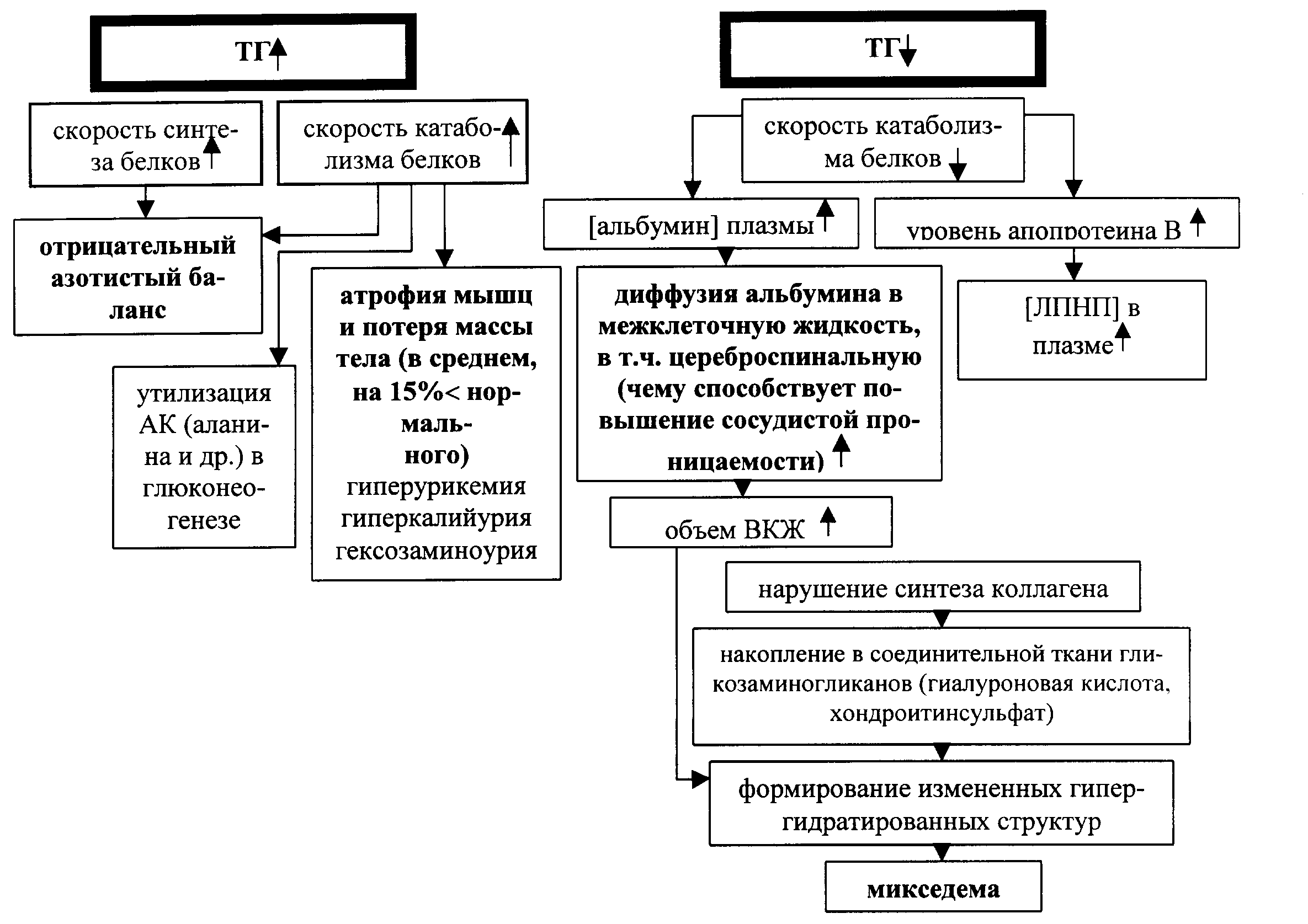
Рис. . Тиреоидные гормоны и белковый обмен
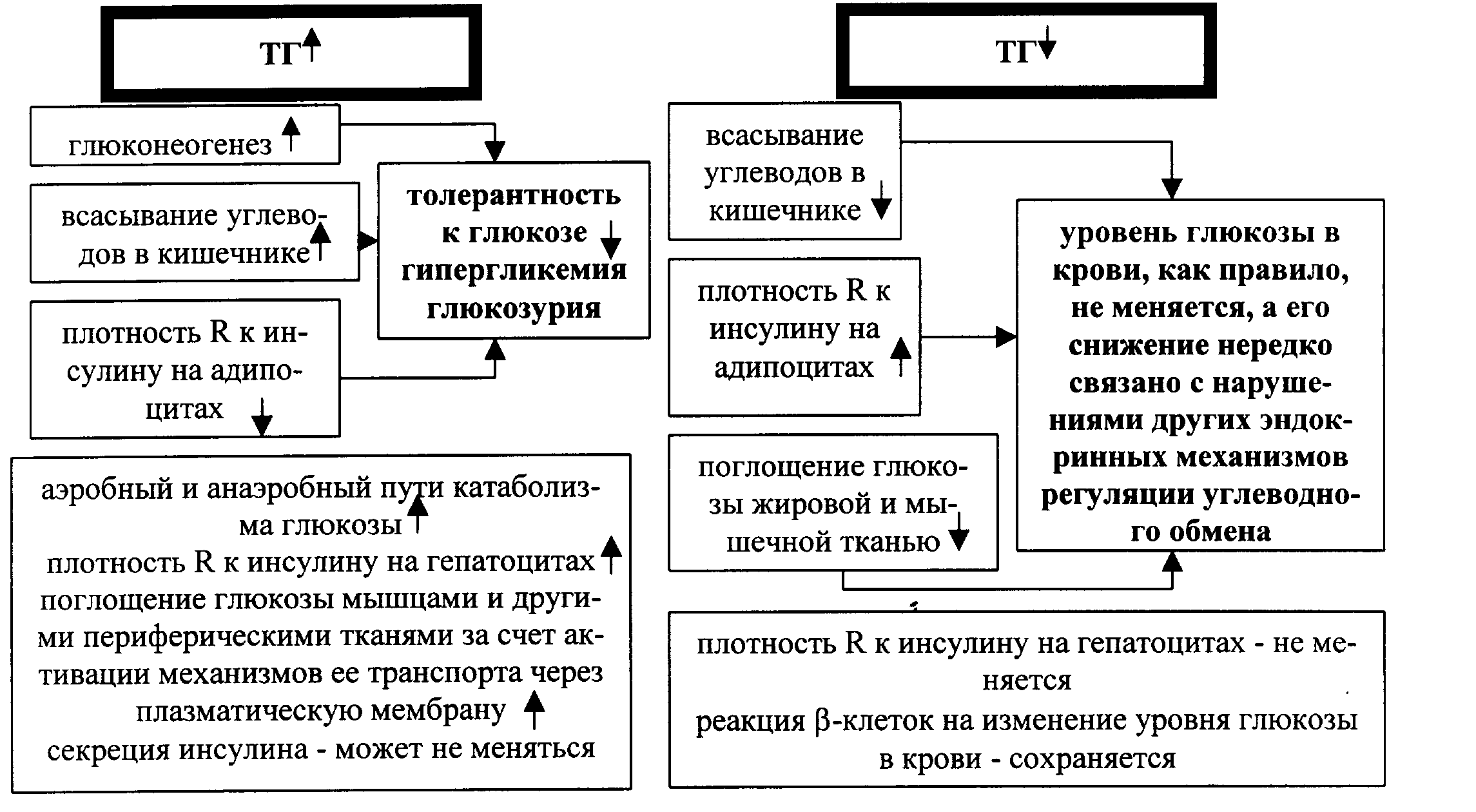
Рис. . Тиреоидные гормоны и углеводный обмен
Влияние тиреоионых гормонов на систему кровообращения реализуется прежде всего их прямыми и опосредованными эффектами на миокард .
Прямое действие йодтиронинов на сердце обеспечивается их влиянием на определенные участки клеточного генома кардиомиоцитов. В результате усиливается экспрессия генов, контролирующих синтез структурных и регуляторных белков, необходимых для мышечного сокращения. Прямое действие приводит к развитию тахикардии ( положительный хронотропный эффект). Вместе с тем, повышение активности других пейсмекерных клеток зачастую обусловливает появление эктопических очагов возбуждения, проявляющихся развитием суправентрикулярных аритмий, главным образом, в виде фибрилляции предсердий.
Опосредованный эффект тиреоидных гормонов ни миокард реализуется через их взаимодействие с катехоламинами.
Продолжительное увеличение работы миокарда при патологически повышенном уровне тиреоидных гормонов (состояние тиреотоксикоза) снижает его резервные возможности, что в конечном счете приводит к изнашиванию сердца и к сердечной недостаточности. Развитию последней способствует кардиомиопатия в результате катаболического эффекта тиреоидных гормонов на белковый обмен, затрагивающего и кардиомиоциты.