

19. 2. Пентозофосфатныи путь дыхательного обмена
Наряду с разложением глюкозы в процессе гликолиза существует другой, не менее распространенный путь — пентозофосфатный, или апотомический. В этом случае глюкоза также подвергается первоначальному фосфорилированию с образованием глюкозо-6-фосфата, затем пути расходятся. Монофосфорный эфир глюкозы подвергается окислению при участии фермента глюкозо-б-фосфат- дегидрогеназы. Коферментом является НАДФ, при этом образуетсяфосфоглюконовая кислота. Процесс происходит следующим образом:
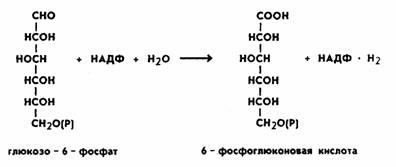
На следующем этапе происходит процесс окислительного декарбоксилирования фосфоглюконовой кислоты. В результате этой peaкции отщепляется одна молекула углекислого газа и образуется пятиуглеродный сахар рибулезо-5-фосфат. Отсюда и название пентозофосфатный цикл или апотомический путь (апотомия— усекновение). Реакция катализируется ферментом фосфоглюконатдегидрогеназой с коферментом НАДФ:
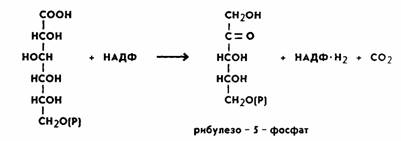
Обе рассмотренные реакции необратимы. Последующие реакции представляют цикл. Для прохождения полного цикла реакций необходимы три молекулыгексозомонофосфата, которые превращаются в три молекулы рибулезо-5-фосфата по уравнению: 3-гексозомонофосфата + 6 НАДФ +ЗН2О →3 пентозофосфата +6 НАДФ-H2 + ЗСО2. Три образовавшиеся молекулы пентозы (рибулезо-5-фосфата) претерпевают превращения. Первая молекула рибулезо-5-фосфата под влиянием фермента эпимеразы превращается в ксилулозо-5 фосфат. Вторая молекула рибулезо-5-фосфата превращается в рибозо-5-фосфат под влиянием ферментафосфопентоизомеразы. Ксилулозо-5-фосфат реагирует с рибозо-5-фосфатом, в результате получается семиуглеродный сахар — седогептулозо-7-фосфат и фосфоглицериновый альдегид. Эта последняя реакция идет под влиянием фермента транскетолазы:

Семиуглеродный сахар реагирует с ФГА, при этом образуется фруктозо-6-фосфат и эритрозофосфат. Фруктозо-6-фосфат изомеризуется до глюкозо-6-фосфата.Эритрозофосфат вместе с третьей молекулой рибулезо-5-фосфата также дает глюкозо-6-фосфат и ФГА. В целом процесс представлен на рисунке 68, а также может быть суммирован в следующем виде:
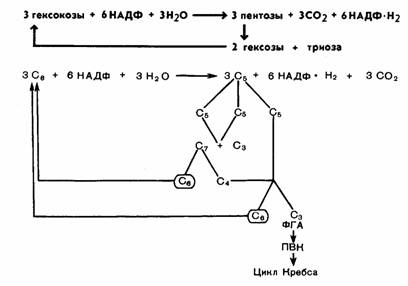
Триоза (ФГА) может претерпевать превращения по гликолитическому пути до пировиноградной кислоты и далее вступать в цикл Кребса. Как видно из уравнения, в пентозофосфатном цикле из каждых трех молекул глюкозы распадается одна, при этом выделяются 3 молекулы СО2 и образуются 6 НАДФ-Н2 и ФГА. Окисление НАДФ-Н2 идет через дыхательную цепь, при этом первоначально НАДФ-Н2 взаимодействует с НАД. При включении 6 НАДФ-Н2 в дыхательную цепь образуется 18 молекул АТФ. Если считать, что 1 молекула АТФ используется на предварительное фосфорилирование гексозы, то накапливается 17 молекул АТФ. Вместе с тем при превращении фосфоглицеринового альдегида в пировиноградную кислоту и ее окислении через цикл Кребса образуется еще 20 молекул АТФ. Таким образом, при окислении по пентозофосфатному пути одной молекулы глюкозы выделяется 37 молекул АТФ. Следовательно, с энергетической стороны апотомический путь почти не отличается от гликолитического. Однако они сильно различаются по промежуточным продуктам. В процессеапотомического распада глюкозы образуется НАДФ-Н2, который может быть использован на различные синтетические процессы, в том числе и в темновыхреакциях фотосинтеза. Важное значение имеет образование пентоз. Пентозы входят в состав нуклеиновых кислот. Нельзя не отметить, что образующийся в этом процессе эритрозофосфт является предшественником лигнина и ряда других соединений Внимательное рассмотрение реакций окисления глюкозы, связанных с пентозофосфатным путем, показывает его большое сходство с превращением углеводов в фотосинтетическом цикле Кальвина Возможно, что благодаря наличию одинаковых промежуточных продуктов эти процессы в ряде пунктов взаимосвязаны. В этой связи интересно отметить, что наряду с локализацией пентозофосфатного пути в цитоплазме он может в отсутствие света осуществляться и в хлоропластах. На протяжении онтогенеза растений и в зависимости от условий происходит смена дыхательных путей. В силу образования различных промежуточных продуктов это оказывает значительное влияние на обмен веществ и физиологическую деятельность растительного организма. Существует ряд методов, с помощью которых можно установить соотношение путей дыхательного обмена (применение специфических ингибиторов, метод с использованием меченых атомов и др.). Однако все они имеют свои недостатки, и в силу этого к экспериментальным данным по этому вопросу надо подходить с большой осторожностью.
ВЛИЯНИЕ ВНЕШНИХ И ВНУТРЕННИХ ФАКТОРОВ НА ИНТЕНСИВНОСТЬ ДЫХАНИЯ
Показатели интенсивности дыхания прямо противоположны показателям интенсивности фотосинтеза. Интенсивность дыхания можно определить: 1) по количеству выделенного СО2; 2) по количеству поглощенного кислорода; 3) по убыли сухой массы. Все эти три показателя рассчитываются на единицу массы в единицу времени