

Частотный оптимум и пессимум ритмической стимуляции
Казалось бы, с увеличением частоты стимулов должна непрерывно увеличиваться частота ответов возбудимых структур, естественно до предела, определяемого лабильностью раздражаемой структуры. Однако на деле ситуация оказывается сложнее.
Для облегчения понимания введём конкретные числовые выражения. Условно определим интервалы временной продолжительности периодов рефрактерности модельной возбудимой структуры:
абсолютная рефрактерность ‑ 5 мс,
относительная рефрактерность – 5 мс.
Отсюда можно рассчитать частоту ритма раздражения, при котором импульсы раздражающего тока будут совпадать с той или иной фазой возбудимости.[++484+C.242]
Расчёт показывает, что при раздражении с частотой ритма менее 100 Гц все пороговые и сверхпороговые стимулы будут восприниматься раздражаемой структурой.
При частоте стимуляции более 100 Гц число воспринятых стимулов в единицу времени будет зависеть от силы стимулов, их способности преодолеть порог раздражения при относительной рефрактерности (рис. 209231646 A).
209231646
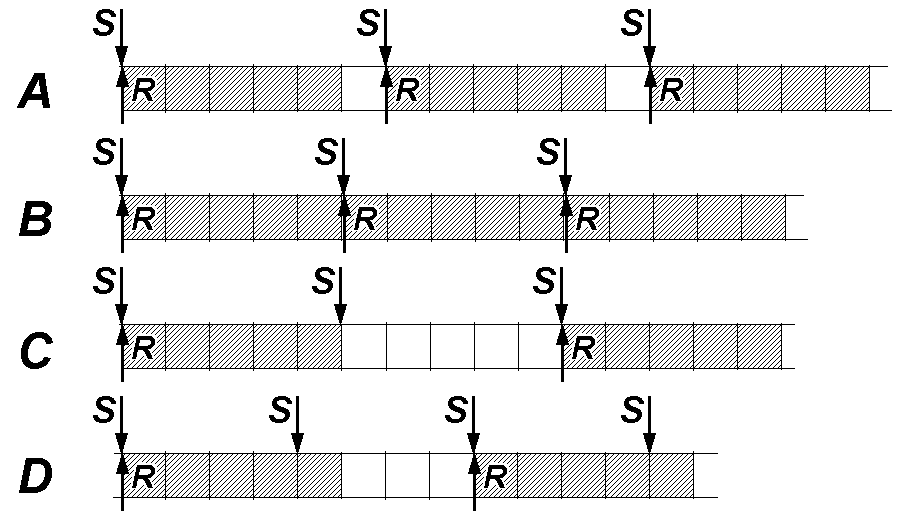
Рис.209231646. Соответствие числа ответов (R) числу стимулов (S). Заштрихованные клеточки соответствуют абсолютной рефрактерности возбудимой структуры. Одна клеточка = 1 мс.
Рассмотрим случай, когда сила стимулов достаточная чтобы преодолеть любой период относительной рефрактерности, тогда препятствием для восприятия стимула будет только абсолютная рефрактерность. Т.е. рефрактерность будет равна 5 мс.
Согласно нашей концептуальной модели раздражение с частотой чуть менее 200 Гц даст максимально возможную частоту ответов равную тем же 200 Гц (рис. 209231646 B).как Как
Следовательно, при раздражении с частотой ритма чуть более 200 Гц многие импульсы тока будут действовать на ткань, когда она не способна отвечать на них. Ритм раздражения подвергается трансформации в более медленный ритм возбуждения. Будет неэффективен каждый 2‑й стимул (рис. 209231646 С). Частота ответов упадёт до 100 Гц.
Дальнейший рост частоты стимуляции приводит к увеличению частоты возбуждений, но каждый второй стимул, а затем третий, четвёртый и т.д. стимул останется без ответа. На рис.209231607 представлена зависимость частоты ответов от частоты стимуляции.
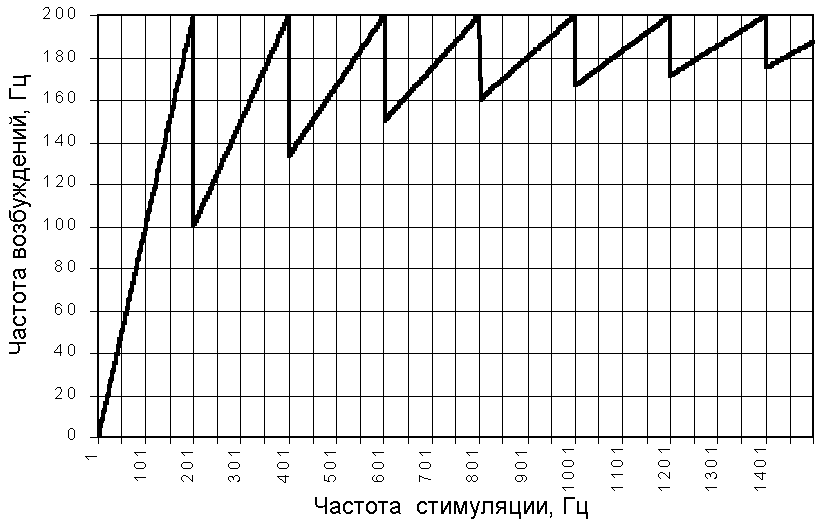
Рис. 209231607. Зависимость частоты ответов возбудимой структуры с рефрактерностью 5 мс (для стимула максимальной силы) от частоты стимуляции.
Как видно из графика, максимальная частота ответов которую мы можем достичь при выбранных параметрах стимула и возбудимой структуры равна 200 Гц.
Частота при которых достигается максимальная частота ответов названы Введенским оптимальной частотой. На графике мы видим, что в нашем примере оптимальными являются частоты кратные 200 Гц — 200, 400, 600, 800 и т.д. Сразу за оптимальными следуют пессимальные частоты, по Н.Е. Введенскому высокие частоты раздражения, вызывающие уменьшение ответа (201, 401, 601, 801 и т. д. Гц).
Как изменятся частотные оптимумы и пессимумы раздражения при снижении силы стимуляции? Очевидно, для этих стимулов возрастёт длительность рефрактерного периода.
Как видно из графика (рис. 209231608), максимальная частота ответов, которую мы можем достичь при выбранных параметрах стимула и возбудимой структуры, равна 125 Гц. Оптимальными являются частоты кратные 125 Гц.
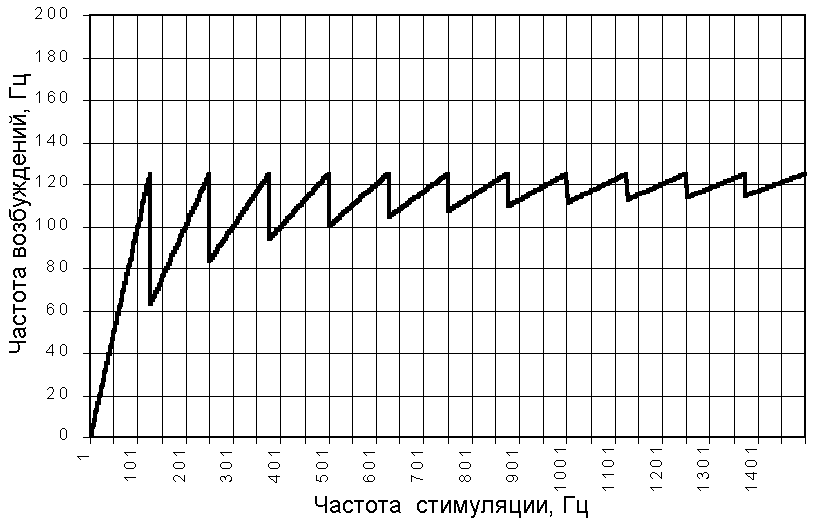
Рис. 209231608. Зависимость частоты ответов возбудимой структуры с рефрактерностью 8 мс (для стимула средней силы) от частоты стимуляции.
При использовании пороговой стимулов рефрактерность возбудимой структуры будет равна АРП+ОРП, в нашем примере 10 мс. Как видно из графика (рис. 209231609), в этом случае максимальная частота ответов, которая достижима при выбранных параметрах стимула и возбудимой структуры равна 100 Гц. И как мы уже отметили, именно этой частоте кратны оптимальные частоты раздражения.
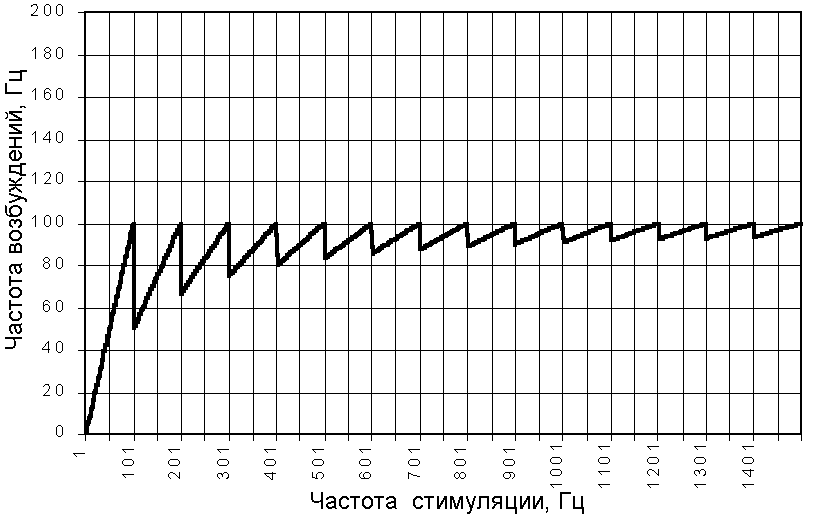
Рис. 209231608. Зависимость частоты ответов возбудимой структуры с рефрактерностью 8 мс (для стимула средней силы) от частоты стимулции.
Можно сделать вывод, что если выполняются законы раздражения (силы, времени, градиента) для одиночного стимула, то оптимум при серийных раздражениях этим стимулом будет не больше 1/АРП и не меньше 1/(АРП+ОРП). Т.е. лабильность определяется длительностью рефрактерных периодов.
Лабильность неодинакова не только у разных тканей, но и у разных структурных единиц одной и той же ткани. Более того, даже у клетки лабильность непостоянна и определяется ее функциональным состоянием.
Не следует путать понятия «усталость», «пессимальное торможение» и пессимальная частота раздражения. Если мы наблюдаем пессимальную частоту раздражения, стоит нам незначительно уменьшить частоту стимуляции и мы отметим значительный рост частоты возбуждений.
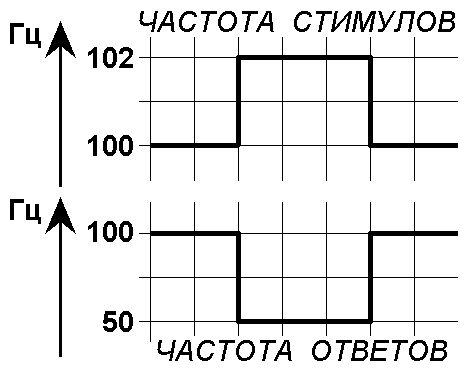
Рис. 209251331
При усталости и пессимальном торможении этого не произойдет. Причину мы выясним в дальнейшем.