

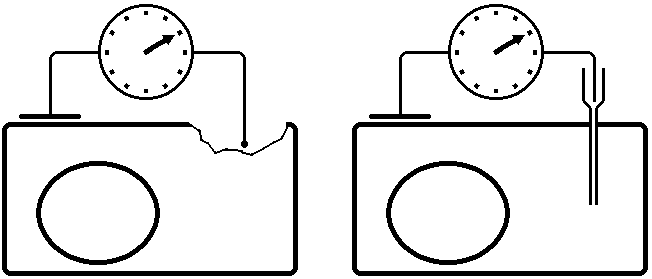
С) Регистрация и измерение потенциала покоя.
Регистрация потенциала покоя
методом повреждения
м
 етодом
внутриклеточного отведения
етодом
внутриклеточного отведения
Внутриклеточный метод регистрации ПП
Микроэлектрод устанавливают над исследуемым объектом, например скелетной мышцей, а затем при помощи микроманипулятора вводят внутрь клетки.
При удачном введении микроэлектрода мембрана плотно охватывает его кончик и клетка сохраняет способность функционировать в течение нескольких часов, не проявляя признаков повреждения.
Микроэлектрод является активным (референтным). Электрод сравнения (индифферентный) обычных размеров погружают в нормальный солевой раствор, в котором находится исследуемая ткань.
Д) Электрогенез потенциала покоя.
Значение потенциала покоя клетки определяется двумя основными факторами:
соотношением концентраций проникающих через покоящуюся поверхностную мембрану катионов и анионов
соотношением проницаемостей мембраны для этих ионов
Для количественного описания этой закономерности используют уравнение Гольдмана - Ходжкина - Kатца:
![]()
где Em - потенциал покоя; R – газовая постоянная; T – абсолютная температура; F – постоянная Фарадея; РK, РNa, РCl - проницаемости мембраны для ионов K+, Na+ и Сl- соответственно; Ko+, Nao+ и Сlo- - наружные концентрации ионов K+, Na+ и Сl-, а Ki+, Nai+ и Сli- - их внутренние концентрации.
Основным механизмом формирования потенциала покоя являются
с
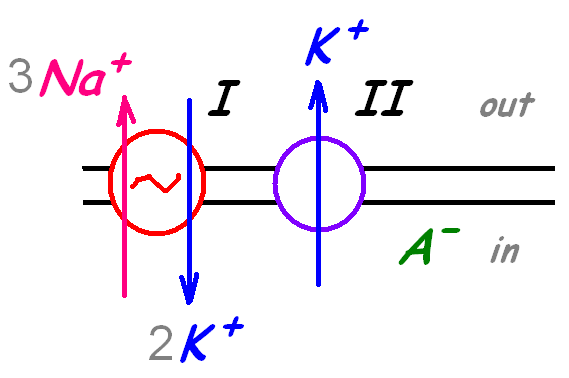 оздание
концентрационной асимметрии K+ при
работе калий-натриевого насоса
(калий-натриевой АТФазы)
оздание
концентрационной асимметрии K+ при
работе калий-натриевого насоса
(калий-натриевой АТФазы)
выход K+ из клетки по градиенту концентрации
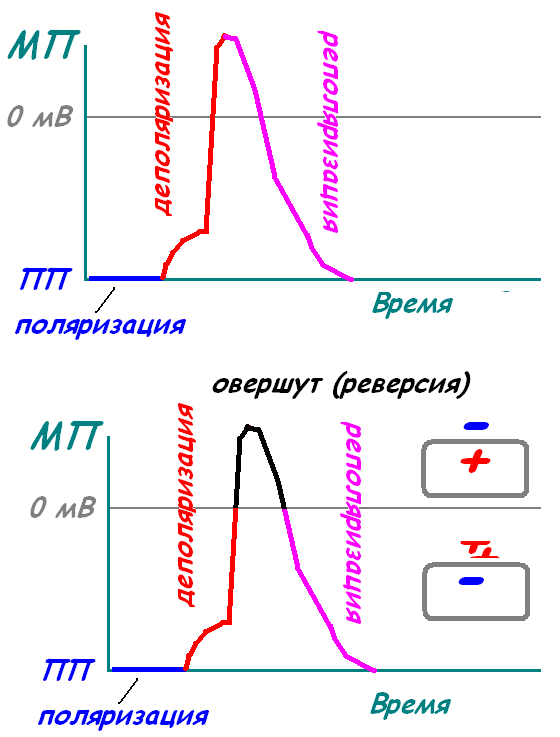
Изменения потенциала покоя
Уменьшение ПП - деполяризация
(ПП становится менее отрицательным)
Увеличение ПП - гиперполяризация
(ПП становится более отрицательным)
-
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ
ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ»)
Кафедра медико-биологических дисциплин
Специальность (направление подготовки)
Курс
второй
060101.65 лечебное дело,
060103.65 педиатрия
Семестр
четвёртый
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № ii (1)
Потенциал действия, его фазы, их происхождение. Соотношение периодов возбудимости с фазами потенциала действия.
Утверждено на заседании кафедры медико-биологических дисциплин,
протокол от 12.05.11 № 10.
Преподаватель к.м.н.
В.Г. Нестеров
Зав. кафедрой д.м.н., профессор
М.И. Чурносов
быстрое колебание мембранного потенциала, возникающее при возбуждении нервных и мышечных клеток (волокон); активный электрический сигнал, с помощью которого осуществляется передача информации в организме человека и животных. Основан на быстро обратимых изменениях ионной проницаемости клеточной мембраны, связанных с активацией и инактивацией ионных мембранных каналов.
ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала, равного -90 мВ.
Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь аксона. Нормальное поляризованное состояние в -90 мВ немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, В крупных нервных волокнах значительный избыток входящих внутрь положительных ионов натрия обычно приводит к тому, что мембранный потенциал «проскакивает» за пределы нулевого уровня, становясь слегка положительным. В некоторых более мелких волокнах, как и в большинстве нейронов центральной нервной системы, потенциал достигает нулевого уровня, не «перескакивая» его.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
Электроупавляемые натриевые и калиевые каналы. Необходимым участником процессов деполяризации и реполяризации во время развития потенциала действия в мембране нервного волокна является электроуправляемый натриевый канал. Электроуправляемый калиевый канал также играет важную роль в увеличении скорости реполяризации мембраны. Оба типа электроуправляемых каналов существуют дополнительно к Na+/K+ -насосу и каналам К*/Na+-утечки.
Электроуправляемый натриевый канал. В верхней части рисунка показан электроуправляемый натриевый канал в трех различных состояниях. Этот канал имеет двое ворот: одни вблизи наружной части канала, которые называют активационными воротами, другие — у внутренней части канала, которые называют инактивационными воротами. В верхней левой части рисунка изображено состояние этих ворот в покое, когда мембранный потенциал покоя равен -90 мВ. В этих условиях активационные ворота закрыты и препятствуют поступлению ионов натрия внутрь волокна.
Активация натриевого канала. Когда мембранный потенциал покоя смещается в направлении менее отрицательных значений, поднимаясь от -90 мВ в сторону нуля, на определенном уровне (обычно между -70 и -50 мВ) происходит внезапное конформационное изменение актива-ционных ворот, в результате они переходят в полностью открытое состояние. Это состояние называют активированным состоянием канала, при котором ионы натрия могут свободно входить через него внутрь волокна; при этом натриевая проницаемость мембраны возрастает в диапазоне от 500 до 5000 раз.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
-
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ
ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ»)
Кафедра медико-биологических дисциплин
Специальность (направление подготовки)
Курс
второй
060101.65 лечебное дело,
060103.65 педиатрия
Семестр
четвёртый
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № iii (1)
Критерии оценки возбудимости тканей. Законы раздражения и возбуждения.
Утверждено на заседании кафедры медико-биологических дисциплин,
протокол от 12.05.11 № 10.
Преподаватель к.м.н.
В.Г. Нестеров
Зав. кафедрой д.м.н., профессор
М.И. Чурносов
Возбудимость клетки меняется не только в процессе ее возбуждения, но и при изменении химического состава внеклеточной жидкости, например, в результате длительной высокой активности клеток, отклонения показателей внутренней среды в патологических случаях. Возбудимость различных нейронов вариабельна. Наиболее возбудимы нейроны ретикулярной формации.
Показателями состояния возбудимости ткани являются пороговый потенциал, пороговая сила и пороговое время.
Характер ответных реакций отдельных возбудимых структур и организма в целом зависит от силы, длительности, скорости нарастания (градиента) силы раздражителя. Определение законов раздражения может быть очень коротким и всех трёх сразу:
Раздражитель может вызвать возбуждение, если он достаточно сильный (закон силы), длительный (закон времени) и быстро нарастает (закон градиента).
Строго говоря эти законы раздражения выполняются для элементарных возбудимых структур (участка мембраны, клетки, волокна) или для ответа минимальной силы составной структуры (мышца, нерв). Закон силы
Прежде всего, необходимо помнить, что возбуждение может возникнуть при деполяризации мембраны до критического уровня (КУД). Раздражитель минимальной силы, вызывающий возбуждение называется пороговым. Раздражитель, сила которого превышает пороговый уровень, называется сверхпороговым. Следует обратить внимание на то, что, чем больше сила сверхпорогового раздражителя, тем быстрее возникает возбуждение.
Введение понятия «порог раздражения» как следствия закона силы является очень важным для оценки возбудимости объекта.
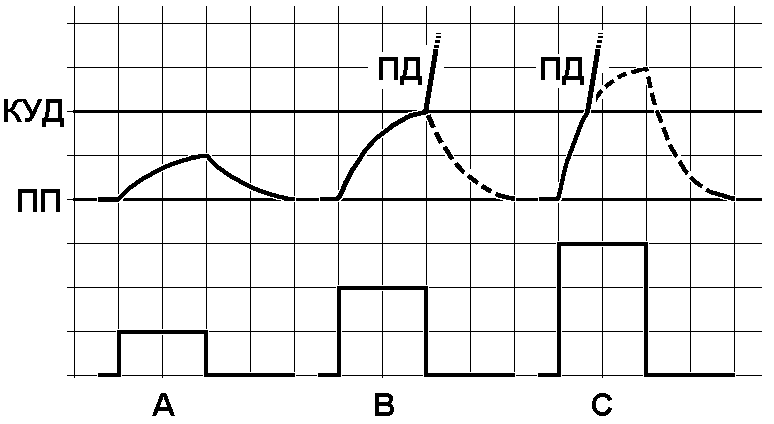 Закон
силы.A ‑
подпороговый стимул, B – пороговый
стимул, C – сверхпороговый
стимул.
Закон
силы.A ‑
подпороговый стимул, B – пороговый
стимул, C – сверхпороговый
стимул.
Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
Закон времени
Следует подчеркнуть, что согласно закону времени, слишком короткие по длительности стимулы не способны вызвать возбуждение, какими бы сильными они небыли. Это используется в физиотерапии при получении калорического эффекта при воздействии токами высокой частоты.
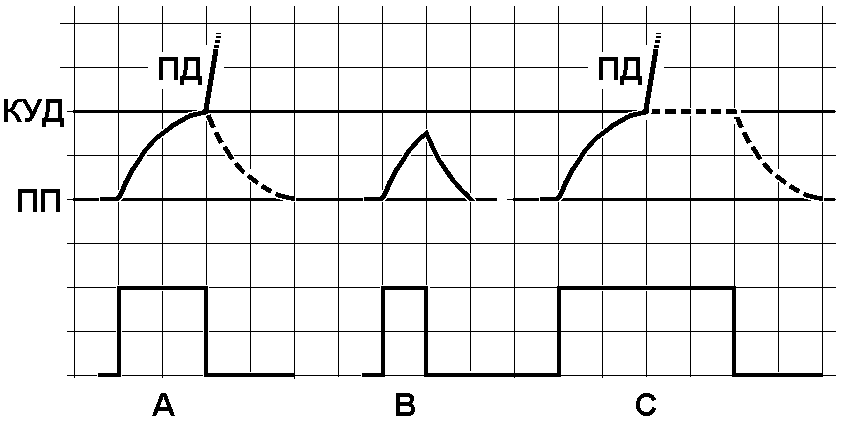
Рис. 209271137. Закон времени
A — подпороговый стимул достаточной длительности для возникновения потенциала действия, B — пороговый стимул, недостаточной длительности для возникновения потенциала действия C — подпороговый стимул более чем достаточной длительности для возникновения потенциала действия.
Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
Важным следствием закона времени является введение понятия полезное время – минимальное время, которое необходимо для действия рассматриваемого раздражителя, чтобы возникло возбуждение. Почему полезное? Потому что дальнейшее действие раздражителя на структуру в состоянии возбуждения бесполезно, ничего уже не изменишь. Бесполезно теряется время.
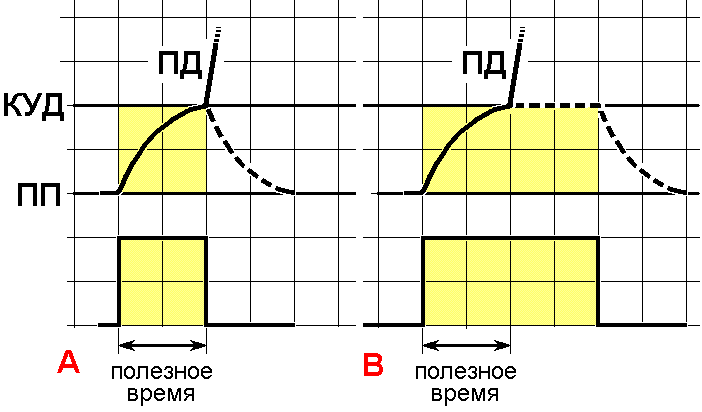 Полезное
время при стимулах разной длительности.
Полезное
время при стимулах разной длительности.
A ‑ стимул по длительности равен полезному времени, B – стимул по длительности больше полезного времени.
Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
Чем больше сила раздражителя, тем меньше полезное время. Но об этом чуть позже.
Закон градиента
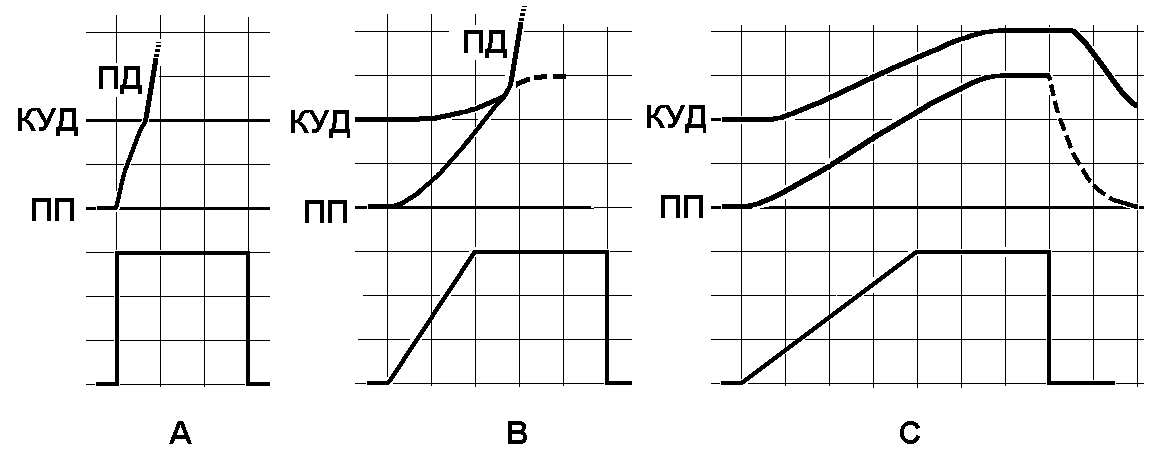
Закон градиента.
A — прямоугольный пороговый стимул, B —стимул, с линейно нарастающей силой, вызывающий ПД, C — стимул, с более полого нарастающей силой, не вызывающий ПД.
Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия, МП – мембранный потенциал. Объяснения в тексте.
Зависимость ответной реакции от градиента раздражения. Зависимость ответной реакции от градиента раздражения была подмечена еще Дюбуа-Реймоном, который определил, что результат действия электрического тока на ткань зависит от быстроты изменения плотности силовых линий раздражающего тока. Градиент раздражения отражает взаимосвязь между ответной реакцией, силой раздражения и временем его воздействия. Например, если взять несколько раздражителей пороговой силы с разным градиентом, то время их действия на клетку, волокно или ткань должно быть тем больше, чем меньше скорость нарастания этих раздражителей до пороговой величины. Раздражители с очень медленной скоростью нарастания, даже если они действуют очень длительное время, вообще не способны вызвать ответную реакцию типа распространяющегося возбуждения (рис. ).
Эти закономерности лежат в основе физиологического явления, получившего название аккомодации. Аккомодация возбудимых структур, которая возникает при воздействии медленно нарастающих по силе раздражителей, выражается в повышении порога возбудимости. Считают, что механизм аккомодации к медленно нарастающему раздражению у различных возбудимых структурных образований организма принципиально одинаков и обусловлен более быстрым и опережающим нарастанием критического уровня деполяризации по сравнению с развитием местных деполяризующих процессов мембраны.