Биохимическая классификация
По биохимической классификации мембранные белки делятся на интегральные и периферические.
Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентов или неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны.
Д) Значение гликокаликса
Собственно надмембранный комплекс, или гликокаликс толщиной 10-20 нм. В его состав входят периферические белки мембраны, углеводные части гликолипидов и гликопротеинов. Гликокаликс играет важную роль в рецепторной функции, обеспечивает «индивидуализацию» клетки — в его составе сосредоточены рецепторы тканевой совместимости.
Гликокаликс
Гликокаликс представляет собой внешний по отношению к клеточной мембране слой.
Он состоит из гликопротеинов, протеогликанов и глюкозаминогликанов и связывается с мембранными структурами с помощью специальных белков-рецепторов, объединяя цитоскелет, мембрану и внеклеточный матрикс в динамическую, подвижную структуру (рис.807251108).
Как показали электронно-микроскопические исследования, гликокаликс имеет вид рыхлого волокнистого слоя толщиной 3-5 нм, покрывающего всю поверхность клетки (рис. 807251142).
В состав гликокаликса входят полисахаридные цепочки мембранных интегральных белков – гликопротеидов. Они содержат такие углеводы, как манноза, глюкоза, сиаловая кислота и др. Углеводные гетерополимеры гликокаликса образуют ветвящиеся цепочки, между которыми располагаются свободные гликолипиды и протеогликаны.
Слой гликокаликса сильно обводнен, имеет желеобразную консистенцию, что значительно снижает диффузию различных веществ из клетки. Здесь же могут накапливаться выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в клетку.
Мембранный потенциал – это разность потенциалов между цитоплазмой и окружающим клетку наружным раствором.
При измерении мембранного потенциала активный электрод располагают внутри клетки, пассивный – снаружи.
Мембранный потенциал покоя (ПП) - мембранный потенциал клетки в состоянии физиологического покоя
У различных клеток мембранный потенциал покоя варьирует от –50 мВ и до –90 мВ
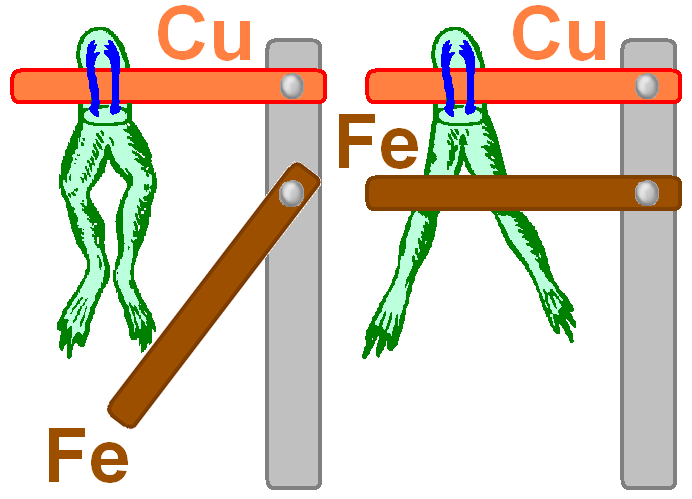
Б) Опыты л.Гальвани.
Начало систематического изучения биоэлектрических явлений связывают с именем итальянского физика и анатома Луиджи Гальвани (Galvani L.).
Л.Гальвани первым убедился в существовании "живого электричества". Термин «животное электричество» принадлежит Л.Гальвани.
Это произошло в 1771 г. По данным других источников — в 1780 г., но опубликованы результаты открытия были только в 1791 г.
Первый («балконный») опыт Л.Гальвани
Учёного интересовало влияние электрических грозовых разрядов на мышцы лягушки.
Препарат задних лапок лягушек на медном крючке был подвешен в грозу к железному балкону.
Влияние молнии на мышцы лягушки он не заметил, но отметил другое — от ветра в дождь препарат задевал балконные перила, и в этот момент мышцы сокращались.
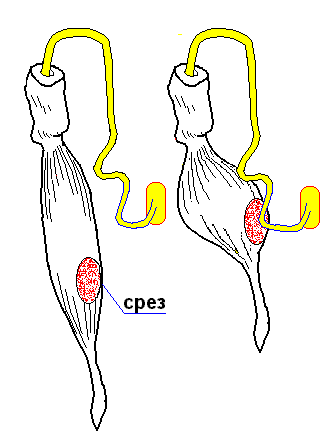
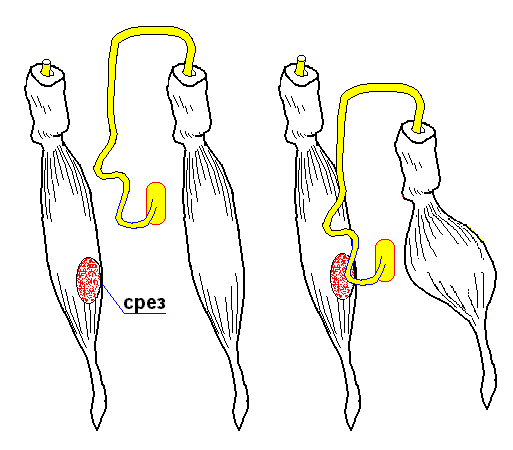
Второй опыт Л.Гальвани (без металла)
Набрасывался нерв между поврежденной (срезом) и неповрежденной поверхностями мышцы
В ответ - сокращение интактной мышцы