

Рецепторы постсинаптической мембраны
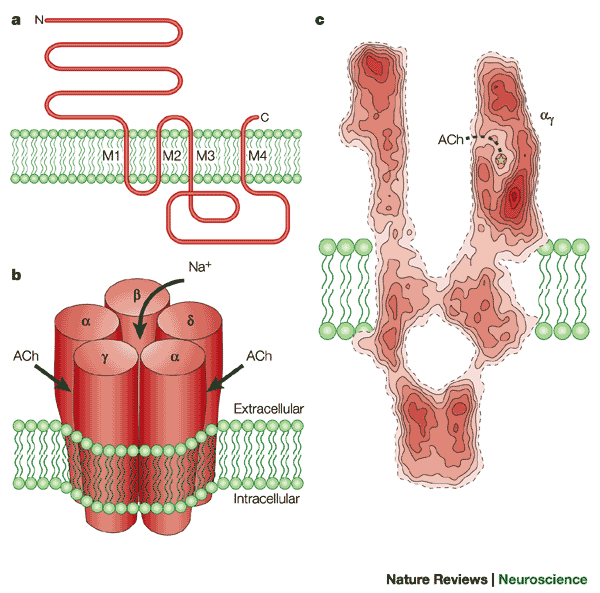
Никотиновый рецепторный канал представляет собой гликопротеин, построенный из пяти субъединиц (рис. 809250501, 809252331).
Рис. 809252331. Никотиновый рецептор, формирующий ионный канал.
|
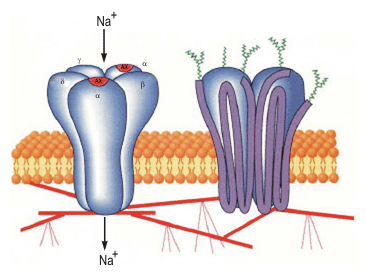
Две -субъединицы содержат два связывающих ацетилхолин сайта, ассоциированных с каналом. При взаимодействии этих сайтов с ацетилхолином значительно увеличивается вероятность открытия канала. Три других субъединицы -субъединица, -субъединица и -субъединица доукомплектовывают протеин. Субъединицы, полипептидные цепи которых четыре раза пронизывают липидный бислой, с внешней стороны гликозилированы, а внутри взаимодействуют с белками тубулинового и актинового цитоскелета.
Электронная микроскопия показывает, что никотиновый рецепторный канал - это симметричная структура, похожая на цилиндр, живой поясок с каналом в центре. Приблизительно по одной трети протеина выступает с обеих сторон плазмалеммы наружу и внутрь клетки, остальная часть находится в толще мембраны. Наружный выступ напоминает воронку, являющуюся входом в канал.
Никотиновые каналы обладают лишь относительной специфичностью. Они обеспечивают диффузию любых катионов. Поскольку покоящаяся мембрана имеет потенциал близкий к равновесному потенциалу для K+, то это означает, что будет диффундировать много больше ионов натрия Na+ и кальция Ca2+ внутрь клетки, чем калия K+ наружу. Это является причиной деполяризации постсинаптической мембраны и возбуждения мышечной клетки. В отличие от других синапсов, этот вид деполяризации получил название потенциала концевой пластинки (ПКП).
Варианты изображения н-холинорецептора (nicotinic acetylcholine receptors)
|
xiv |
http://www.nature.com/nrn/journal
|
Этапы передачи возбуждения в НМС
|
Ацетилхолин в синаптической щели быстро разрушается ацетилхолинэстеразой (АХЭ), превращаясь в холин (он вновь захватывается для последующего синтеза) и ацетат.
|
Миниатюрный потенциал концевой пластинки
|
xvi |
Один квант ацетилхолина активирует ~ 1,5·103 каналов. Через один активированный канал нейромышечного соединения проходит приблизительно 2·104 одновалентных ионов. Постоянная времени затухания миниатюрного потенциала концевой пластинки соответствует постоянной времени закрытия канала. Постоянная времени закрытия канала зависит от разности потенциалов на мембране (потенциалзависимый канал). Деполяризация продлевает, а гиперполяризация укорачивает время открытых каналов.
Фармакологическая модификации нмс
|
xvii |
Как и все синапсы, нервно-мышечный синапс подвергается фармакологической модификации (см. подробно в учебнике [1] С.71).
Коротко. Можно блокировать проведение ПД по пресинаптическому элементу (новокаином), блокировать высвобождение медиатора, например, за счет удаления из среды ионов кальция или добавления ионов марганца, или при помощи ботулинического токсина, можно заблокировать синтез АХ, угнетая захват холина. Наконец, что в практическом отношении чрезвычайно важно, можно блокировать сами рецепторы и, тем самым, полностью прекратить передачу возбуждения в синапсе. Это можно сделать, используя такие вещества, которые при высоких концентрациях способны вытеснять ацетилхолин с ХР. Этим свойством обладают кураре и курареподобные вещества (д‑тубокурарин, диплацин и т.д.). Эта процедура находит широкое применение в хирургии. Существует также возможность управлять активностью ацетилхолинэстеразы (АХЭ). Если ее активность уменьшить до определенной степени, то это будет способствовать накоплению АХ в синаптической щели. Такая ситуация при определенной патологии (миастения гравис, когда количество выбрасываемого медиатора резко снижено) является благоприятной. Если инактивация АХЭ достигает значительной величины, то это сопровождается развитием стойкой деполяризации в области синапса и приводит к блокаде проведения возбуждения через мионевральный синапс. В конечном итоге это может привести к гибели организма. На этом явлении основано применение фосфороорганических соединений (ФОС) в качестве отравляющих веществ (ОВ) или в качестве инсектицидов (дихлофос, хлорофос и т.д.).
Рецептор. Рецептивное поле и рефлексогенная зона.
Рефлексогенная зона (син. рецептивное поле) — область тела (например, участок кожи, слизистой оболочки, сосудистой стенки), в пределах которой расположены рецепторы одного типа, раздражение которых приводит к возникновению определенного рефлекса.
Рефлексогенные зоны, рецептивные поля рефлексов, области расположения специальных рецепторов в организме, раздражение которых вызывает те или др. специфические безусловные рефлексы. Например, раздражение слизистой оболочки носоглотки вызывает рефлекс чихания, а слизистой оболочки трахеи, бронхов — рефлекс кашля. Р. з., находящиеся на поверхности кожи, могут перекрывать одна другую, вследствие чего раздражение, наносимое на определённый участок кожи, в зависимости от его силы и состояния центральной нервной системы вызывает то один, то другой рефлекс. Сложно организованные Р. з. находятся во внутренних органах и участвуют в рефлекторной регуляции их функциональной деятельности. Например, при раздражении барорецепторов дуги аорты и каротидного синуса возникают депрессорные рефлексы — снижение кровяного давления и урежение сердцебиений. Рефлекторный ответ на раздражение Р. з. может иногда тормозиться под влиянием других нервных центров, не имеющих отношения к данному рефлексу (см. Доминанта). Р. з. относятся к структурной организации одних только безусловных рефлексов, так как условные рефлексы не имеют стабильных рефлекторных дуг. Формирование Р. з. определяется наследственной организацией нервной системы и её созреванием в процессе индивидуального развития организма.
П. А. Киселёв.
Рецептивное поле (от лат. receptus – принятый, полученный) — функциональная объединение рецепторов, информация от которых поступает в единый центр. Таким центром может выступать суммирующая нервная клетка, собирающая импульсы непосредственно от рецепторов или от рецептивных полей.
РЕЦЕПТИВНОЕ ПОЛЕ (от лат. receptus — принятый, полученный) — функциональная единица сенсорной системы, которая осуществляет переработку информации, полученной рецепторами. Морфофизиологическое Р. п. представляет собой совокупность рецепторов, соединенных с нервной клеткой, которая называется выходной (или суммирующей). Р. п. делятся на простые и сложные.
Простое Р. п. — это совокупность нескольких рецепторов, контактирующих с одной нервной клеткой. Сложное Р. п. — это совокупность нескольких простых рецептивных полей, образованная путем контактирования выходных нервных клеток между собой возбуждающими или тормозными синапсами. В результате переработки информации в Р. п. выделяются основные характеристики раздражителей.
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования |
|||
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ») |
|||
Кафедра медико-биологических дисциплин
|
|||
|
|||
Специальность (направление подготовки) |
Курс |
второй |
|
060101.65 лечебное дело, 060103.65 педиатрия |
Семестр |
четвёртый |
|
|
|
|
|
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ |
|||
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № xviii (1) |
|||
|
|||
Утверждено на заседании кафедры медико-биологических дисциплин, протокол от 12.05.11 № 10. |
|||
|
|||
|
|
||
|
|
||
Скелетная мышца состоит из мышечных волокон. У человека количество этих волокон в мышце устанавливается через 4 ‑ 5 месяцев после рождения и изменяться под воздействием тренировки.
Мышечное волокно покрыто тонкой эластичной мембраной — сарколеммой. Ее структура подобна структуре мембран других клеток, в частности нервных. Мембрана мышечных клеток играет важную роль в возникновении и проведении возбуждения.
Внутреннее содержимое мышечного волокна называется саркоплазмой. Она состоит из двух частей. Первая — саркоплазматический матрикс — представляет собой жидкость, в которую погружены сократительные элементы мышечного волокна — миофибриллы. В этой жидкости находятся растворимые белки (например, миоглобин), гранулы гликогена, капельки жира, фосфатсодержащие вещества и другие малые молекулы и ионы.
Вторая часть саркоплазмы — саркоплазматический ретикулум. Так обозначается система сложносвязанных между собой элементов в виде вытянутых мешочков и продольных трубочек, расположенных между миофибриллами параллельно им. Мышечное волокно внутри пересекают поперечные Т-трубочки (Т-система). Выстилающие их мембраны по своей структуре сходны с сарколеммой. Поперечные трубочки соединяются с поверхностной мембраной мышечного волокна, связывая его внутренние части с межклеточным пространством. Продольные трубочки примыкают к поперечным, образуя в зоне контактов так называемые цистерны. Эти цистерны отделены от поперечных трубочек узкой щелью. На продольном разрезе волокна видна характерная структура — триада, включающая поперечную трубочку с прилегающими к ней с двух сторон цистернами. Ретикулярные триады фиксированы так, что их центр находится вблизи границы A‑ и I‑дисков (см. ниже).
Саркоплазматический ретикулум играет важную роль в передаче возбуждения от поверхностной мембраны волокна вглубь к миофибриллами в акте сокращения. Через cаркоплазматический ретикулум и поперечные трубочки может также происходить выделение продуктов обмена (в частности, молочной кислоты) из мышечной клетки в межклеточное пространство и далее в кровь.
В мышечном волокне содержится до 1000 и более миофибрилл. Каждая из них имеет диаметр 1—3 мкм.
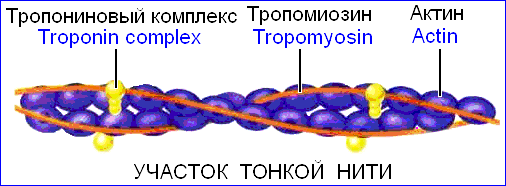
Миофибрилла — это пучок параллельно лежащих нитей (миофиламентов) двух типов — толстых и тонких. Толстые нити состоят из миозина, а тонкие — из актина. Кроме того, в состав тонких миофиламентов входят еще два белка — тропомиозин и тропонин, играющие регуляторную роль в процессах сокращения и расслабления.
Толстый миофиламент образован удлиненными глобулярными молекулами миозина, длинные «хвосты» которых состоят из легкого меромиозина (М-миозина) и собраны в пучок (рис. 709261107).
Рис. 709261107. Толстый миофиламент.
|
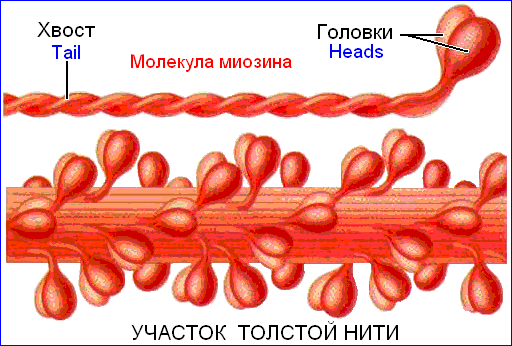
Остальные около 1/6 длины молекулы миозина составляет её «головка», образованная тяжелым меромиозином (Т‑миозином). Он обладает ферментативной АТФ-азной активностью, т.е. способностью катализировать гидролиз АТФ с образованием энергии.
Головки миозиновых молекул повернуты в направлении к тонким миофиламентам и называются поперечными мостиками. По обе стороны от середины толстого миофиламента «головки» молекул миозина повернуты в противоположные стороны, так что средняя часть толстых миофиламентов не имеет поперечных мостиков.
Тонкий миофиламент образован двумя актиновыми нитями, обвитыми одна вокруг другой в виде двойной спирали (рис. 210201735).
Рис. 210201735. Тонкий миофиламент.
|
Механизм мышечного сокращения объясняется моделью скользящих нитей, авторами которой принято считать Х.Хаксли и Дж.Хансона 1 (1954 г.).
Отсюда был сделан вывод, что тонкие и толстые нити при сокращении не изменяют значимо свою длину, а скользят относительно друг друга
-
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ
ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ»)
Кафедра медико-биологических дисциплин
Специальность (направление подготовки)
Курс
второй
060101.65 лечебное дело,
060103.65 педиатрия
Семестр
четвёртый
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № xix (1)
Механизм мышечного сокращения и расслабления.
Биоэлектрические, химические и тепловые процессы в мышцах.
Утверждено на заседании кафедры медико-биологических дисциплин,
протокол от 12.05.11 № 10.
Преподаватель к.м.н.
В.Г. Нестеров
Зав. кафедрой д.м.н., профессор
М.И. Чурносов
Образование потенциала концевой пластинки (ПКП).
Электротоническое распространение ПКП в околосинаптическую область.
Генерация потенциала действия мышечного волокна в околосинаптической области.
Распространение потенциала действия по поверхностной мембране мышечного волокна
Распространение потенциала действия по мембранам поперечных трубочек (Т‑системы).
Реакция рецепторов дигидропиридина 2 на изменение мембранного потенциала.
Передача сигнала дигидропиридиновыми рецепторами рианодиновым рецепторам терминальных цистерн саркоплазматического ретикулума.
Открытие каналов Ca2+, с рианодиновыми рецепторами и выход Ca2+ из саркоплазматического ретикулюма (цистерн) в саркоплазму (цитозоль).
Диффузия Ca2+ в межфибриллярное пространство.
Связывание Ca2+ с тропонином, расположенным на тонком, актиновом, миофиламенте.
Конформационные изменения тропонина при связывании Са2+ «заталкивание» тропомиозина в канавки актиновых нитей (рис. 210201735).
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования |
|||
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ») |
|||
Кафедра медико-биологических дисциплин
|
|||
|
|||
Специальность (направление подготовки) |
Курс |
второй |
|
060101.65 лечебное дело, 060103.65 педиатрия |
Семестр |
четвёртый |
|
|
|
|
|
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ |
|||
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № xx (1) |
|||
|
|||
Анатомо-физиологические свойства гладких мышц. |
|||
Утверждено на заседании кафедры медико-биологических дисциплин, протокол от 12.05.11 № 10. |
|||
|
|||
Преподаватель к.м.н. |
В.Г. Нестеров |
||
Зав. кафедрой д.м.н., профессор |
М.И. Чурносов |
||
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования |
|||
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ») |
|||
Кафедра медико-биологических дисциплин
|
|||
|
|||
Специальность (направление подготовки) |
Курс |
второй |
|
060101.65 лечебное дело, 060103.65 педиатрия |
Семестр |
четвёртый |
|
|
|
|
|
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ |
|||
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № xxi (2) |
|||
|
|||
Понятие о внутренней среде организма: кровь, лимфа, внесосудистые жидкости. |
|||
Утверждено на заседании кафедры медико-биологических дисциплин, протокол от 12.05.11 № 10. |
|||
|
|||
Преподаватель к.м.н. |
В.Г. Нестеров |
||
Зав. кафедрой д.м.н., профессор |
М.И. Чурносов |
||
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования |
|||
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ») |
|||
Кафедра медико-биологических дисциплин
|
|||
|
|||
Специальность (направление подготовки) |
Курс |
второй |
|
060101.65 лечебное дело, 060103.65 педиатрия |
Семестр |
четвёртый |
|
|
|
|
|
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ |
|||
ЭКЗАМЕНА
ЦИОННЫЙ ВОПРОС № (1) |
|||
|
|||
Физиология железистой ткани. |
|||
Утверждено на заседании кафедры медико-биологических дисциплин, протокол от 12.05.11 № 10. |
|||
|
|||
Преподаватель к.м.н. |
В.Г. Нестеров |
||
Зав. кафедрой д.м.н., профессор |
М.И. Чурносов |
||
Внутренняя среда организма – совокупность жидкостей принимающих непосредственное участие в процессах обмена веществ и поддержании гомеостаза организма. 3
Понятие внутренняя среда организма ввел в физиологию К.Бернар в 1854-1857 гг. 4
Внутренняя среда характеризуется динамическим постоянством.
Для описания этого состояния в 1929 г. У.Кэннон ввел термин гомеостаз 5.
В связи с выявлением роли биоритмов в деятельности живого организма хронобиология стала оперировать термином не «гомеостаз», а «гомеокинез» или «гомеорез» , под которым понимается не только значение параметров, но и процесс их изменения во времени.xxii
Однако в литературе чаще используется термин «гомеостаз», при этом имеют ввиду, что постоянство внутренней среды относительно.
Границы гомеостаза могут быть жесткими и пластичными. Их показатели зависят от видовых, индивидуальных, половых и других условий. Жесткими константами являются параметры внутренней среды, которые определяют оптимальную активность ферментов, т.е. возможность осуществления обменных процессов.--162- C.13]