

Структура нервно-мышечного синапса
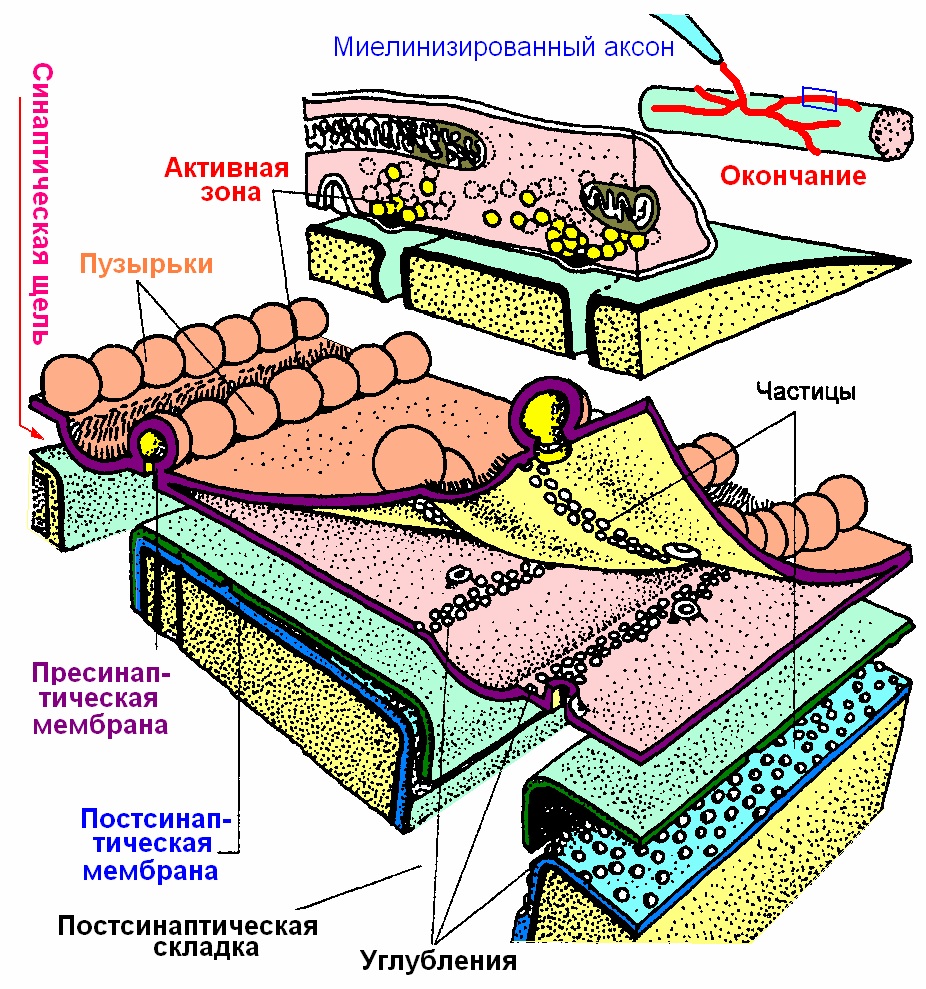
Структура нервно-мышечного синапса в трехмерном изображении показана на рис. 809252134.
Рис. 809252134. Структура нервно-мышечного синапса.
|
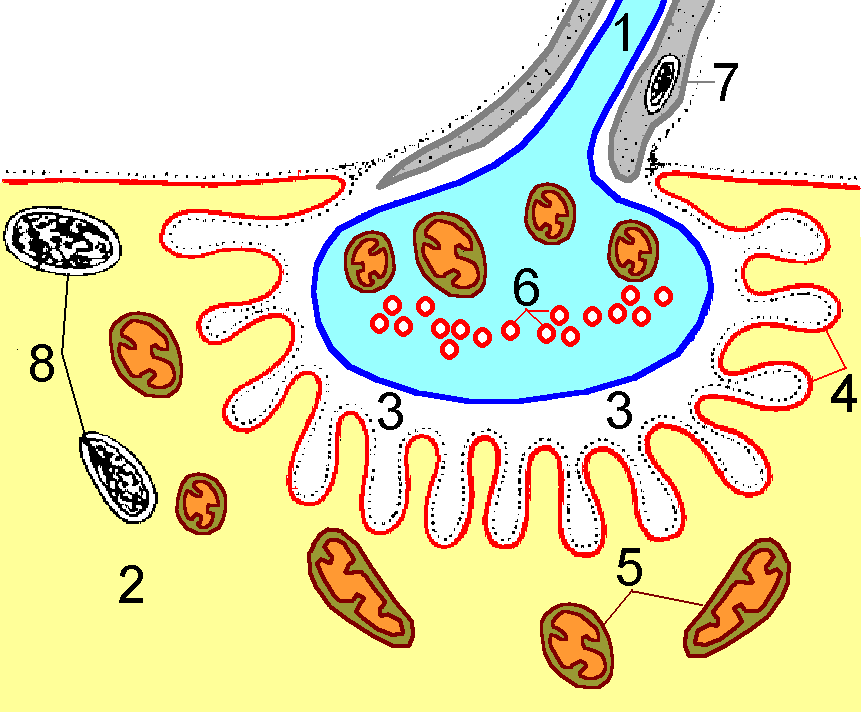
Структура нервно-мышечного синапса в двухмерном изображении, которую легко воспроизвести Вам при ответе, показана на рис. 809252137.
Рис. 809252137. Мионевральный синапс (по Е.А.Шубниковой с изменениями). 1 — аксон мотонейрона: 2 — мышечное волокно; 3 — синаптическая щель; 4 — пальцевидные инвагинации сарколеммы; 5 — митохондрии мышечного волокна; 6 — синаптические пузырьки; 7 — леммоцит; 8 — ядра мышечного волокна.
|

Для НМС характерно наличие большого числа изгибов на пресинаптической и, особенно, на постсинаптической мембране. Благодаря этому, вероятно, возрастает площадь контакта пресинапса с постсинапсом, что увеличивает вероятность взаимодействия.
Деполяризация пресинаптической мембраны пришедшим по аксону потенциалом действия (ПД).
Открытие потенциалзависимых Сa++‑каналов на пресинаптической мембране и поступление Сa++ в пресинапс (пассивный транспорт).
Выход в синаптическую щель ацетилхолина путем экзоцитоза.
Диффузия медиатора к постсинаптической мембране.
Взаимодействие ацетилхолина с Н‑холинорецепторами постсинаптической мембраны мышечного волокна.
Открытие никотиновых рецепторных каналов постсинаптических каналов, пассивный вход Na+ в мышечное волокно.
Образование потенциала концевой пластинки - ПКП (возбуждающего постсинаптического потенциала - ВПСП) в области постсинаптической мембраны.
Электротоническое распространение ПКП в околосинаптическую область.
Формирование ПД мышечного волокна на сарколемме околосинаптической области.
Терминали аксонов содержат большое количество митохондрий и синаптических пузырьков с медиатором (ацетилхолином).
В основном везикулы расположены в определенных местах пресинапса - около так называемых активных зон. В норме везикулы не подходят близко к пресинаптической мембране, вероятно, из-за того, что имеют такой же заряд, как и пресинапс, хотя возможно и наличие жесткой структуры, которая удерживает везикулы в подвешенном состоянии. Каждая везикулам содержит примерно по 1000-10000 молекул ацетилхолина.
Синаптическая мембрана аксона и постсинаптическая мембрана мышечного волокна разделены синаптической щелью.
В синаптической щели расположен гликокаликс - волокна, которые выполняют опорную функцию (связь механическая). Здесь расположена ацетилхолинэстераза (АХЭ), способная расщеплять ацетилхолин со скоростью 1 мл/мс.
В области синапса мышечное волокно не имеет поперечной исчерченности, характерно скопление митохондрий и ядер.
Гребешки постсинаптической мембраны следуют с интервалом примерно в 1 мкм. На вершине гребешка концентрация холинорецепторов достигает максимальных значений (примерно 20000 рецепторов на 1 мкм кв.), а в устьях - т.е. в глубине - около 1000 рецепторов. Во внесинаптической зоне, естественно, концентрация холинорецепторов меньше - порядка 50 на 1 мкм кв.
Молекулярная масса рецепторов нервно-мышечного синапса определена с помощью методики связывания рецептора бунгаротоксином (яд полосатой крайоты - вид змеи) и равна 250.000. Молекула рецептора состоит из 5 субъединиц – 2‑х α, β, γ, δ. Узнающая субъединица - это α. Внешне рецептор похож на гриб. Внутри рецептора проходит ионный канал, пропускающий Na+.