

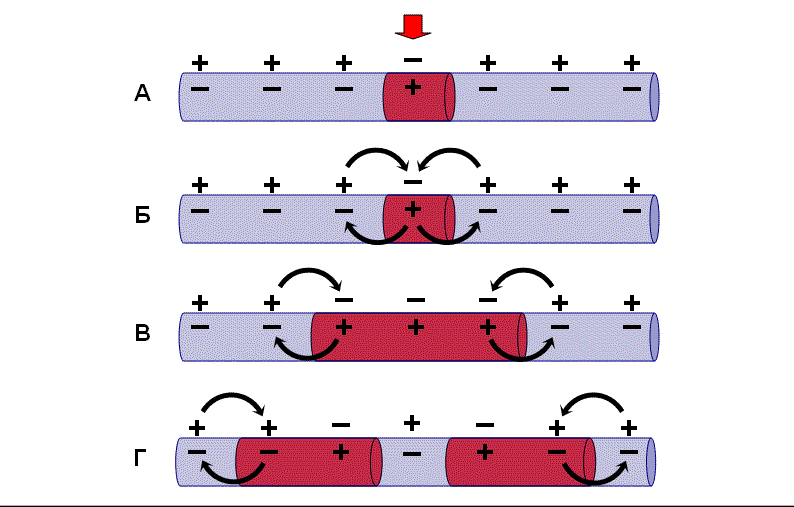
Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток. Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя. Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
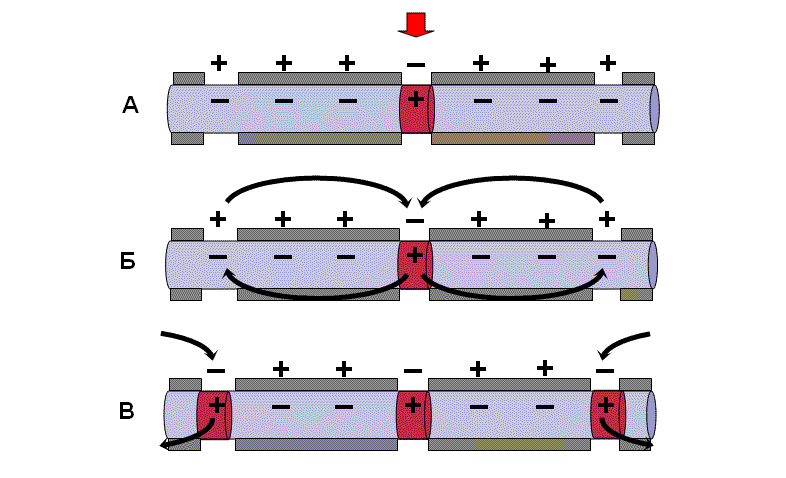
Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны. Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны. Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье. Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
1939 г. американские физиологи
Эрлангер и Гассер
зарегистрировали суммационные потенциалы от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрод
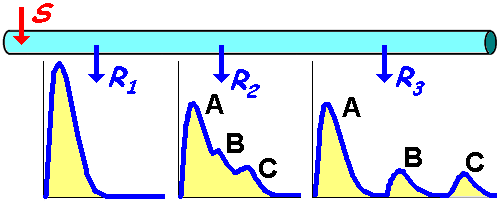
Было установлено, что
чем дальше от места нанесения стимула находился регистрирующий электрод, тем более четко суммарный потенциал делился на ряд пиков,
которые были обозначены буквами латинского алфавита A, B, C.
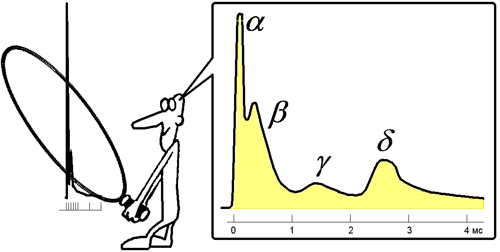
При относительно быстрой скорости записи потенциалов, при более тщательном исследовании и рассмотрении пика A
выделили дополнительно пики и пометили их буквами α (альфа), β (бета), γ (гамма), δ (дельта).
Нервные волокна, составляющие нерв, имеют разную скорость проведения возбуждения.
Нервные волокна можно объединить в группы с примерно одинаковой скоростью проведения возбуждения.
Группы волокон (по Эрлангеру и Гассеру) |
Диаметр, мкм |
Скорость проведения, м/с |
Aa |
13 - 22 |
70 - 120 |
Ab |
8 -13 |
40 - 70 |
Ag |
4 - 8 |
15 – 40 |
Ad |
1 – 4 |
5 – 15 |
B |
1 - 3 |
3 – 14 |
C |
0,5 – 1,0 |
0,5 - 2 |
Нервы у позвоночных состоят из трех основных групп волокон (А, В и С), различающихся по скорости проведения возбуждения, степени миелизации, диаметру волокна, скорости развития ПД.
Значения всех этих показателей в ряду от Aα до С снижаются.
По немиелинизированному волокну ПД распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший ПД за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые ПД. Сами ПД не перемещаются, они исчезают там же, где возникают. Главную роль в возникновении нового ПД играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то ПД будет распространяться в обоих направлениях. Обычно же ПД распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник ПД. Одностороннее проведение ПД обеспечивается свойствами натриевых каналов — после открывания они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» ПД, он не возникает.
При прочих равных условиях распространение ПД по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара ПД может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
ФЕДЕРАЛЬНОЕ Государственное АВТОНОМНОЕ образовательное учреждение высшего профессионального образования |
|||
«БЕЛГОРОДСКИЙ ГОСУДАРСТВЕННЫЙ НАЦИОНАЛЬНЫЙ ИССЛЕДОВАТЕЛЬСКИЙ УНИВЕРСИТЕТ» (НИУ «БелГУ») |
|||
Кафедра медико-биологических дисциплин
|
|||
|
|||
Специальность (направление подготовки) |
Курс |
второй |
|
060101.65 лечебное дело, 060103.65 педиатрия |
Семестр |
четвёртый |
|
|
|
|
|
Дисциплина НОРМАЛЬНАЯ ФИЗИОЛОГИЯ |
|||
ЭКЗАМЕНАЦИОННЫЙ ВОПРОС № vii (1) |
|||
|
|||
Синапсы: строение и классификация. Механизм передачи возбуждения в синапсах. |
|||
Утверждено на заседании кафедры медико-биологических дисциплин, протокол от 12.05.11 № 10. |
|||
|
|||
Преподаватель к.м.н. |
В.Г. Нестеров |
||
Зав. кафедрой д.м.н., профессор |
М.И. Чурносов |
||
Синапс – специализированная зона контакта между нейронами или нейронами и другими клетками, обеспечивающая передачу возбуждения.
Синапсы можно разделить, положив в основу различные критерии.
По характеру контактирующих клеток
выделяют:
Нейрон нейрон (межнейронный, нейронейрональный,)
Нейрон эффекторная клетка, а именно:
нейрон миоцит (нервномышечный, мионевральный)
нейрон гландулоцит (нейросекреторный)
Рецепторная клетка вторичночувствующего рецептора нейрон
По способу передачи сигнала выделяют
химические,
электрические,
смешанные.
Смешанные синапсы называют ещё синапсами двоякого действия.
По конечному эффекту
выделяют синапсы возбуждающие тормозные
По форме контакта
различают (рис. 410020742)
терминальные (концевые, колбообразное соединение);
проходящие (касательные, варикозное расширение аксона).
А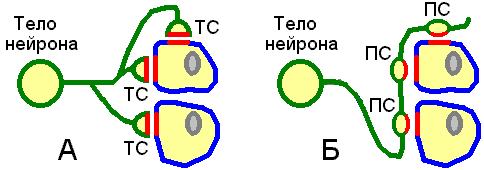 — терминальные
синапсы (ТС); Б — проходящие синапсы
(ПС).
— терминальные
синапсы (ТС); Б — проходящие синапсы
(ПС).
По развитию в онтогенезе
различают
стабильные (например, синапсы дуг безусловного рефлекса)
динамичные (появляющиеся и исчезают в процессе развития организма). Пример — синапсы дуг условного рефлекса.
Типы межнейрональных синапсов
|
 Мозг
человека содержит » 100 миллиардов
нейронов.
Мозг
человека содержит » 100 миллиардов
нейронов.
По характеру контактирующих частей нейронов
выделяют (рис. 410020748):
аксо-аксональные,
аксо-дендритические (дендритные),
аксо-соматические,
дендро-дендритические,
дендро-соматические
и т.п..
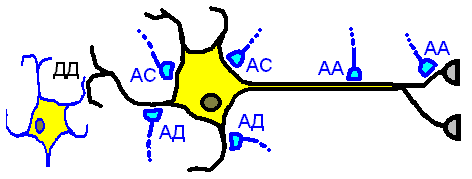
Типы межнейрональных синапсов по характеру контактирующих частей неронов, АД — аксо-дентритические, АС — аксо‑соматические, АА — аксо-аксонаьные, ДД — дендро‑дендритические.
В приложении 1 межнейрональные синапсы показаны более детально.
По локализации в нервной системе
выделяют
центральные (расположены в головном и спинном мозге, т.е. в ЦНС)
периферические (в периферической нервной системе).
Типы химических синапсов
По природе медиатора химические синапсы различают:
холинергические (медиатор - ацетилхолин),
адренергические (медиатор - норадреналин),
дофаминергические (медиатор - дофамин),
ГАМК-ергические (медиатор - гамма-аминомасляная кислота),
глицинергические,
глутаматергические,
аспартатергические,
пептидергические (медиатор - пептиды),
пуринергические (медиатор - АТФ
По характеру рецепторов на постсинаптической мембране, определяющих в постсинапсе процессы, происходящие при передаче возбуждения химические синапсы разделяются на
ионотропные
метаботропные
По характеру изменения мембранного потенциала на постсинаптической мембране при передаче возбуждения химические синапсы разделяются на
возбуждающие
тормозные
Возбуждающими и тормозными могут быть и электрические синапсы. Но у человека тормозных электрических синапсов не найдено, а функция возбуждающих и тормозных химических синапсов у человека хорошо изучена.
Общий план строения синапсов.
С) Проведение возбуждения
Проведение возбуждения через синапс.
Деполяризация пресинаптической мембраны приходящим по аксону потенциалом действия (ПД).
|
Этапы передачи сигнала в химическом синапсе. При огромном разнообразии синапсов и различиях в механизмах передачи сигнала в них, для большинства изученных синапсов можно выделить следующие наиболее общие этапы передачи возбуждения:
Везикулярная гипотеза освобождения медиатора в синапсе
|
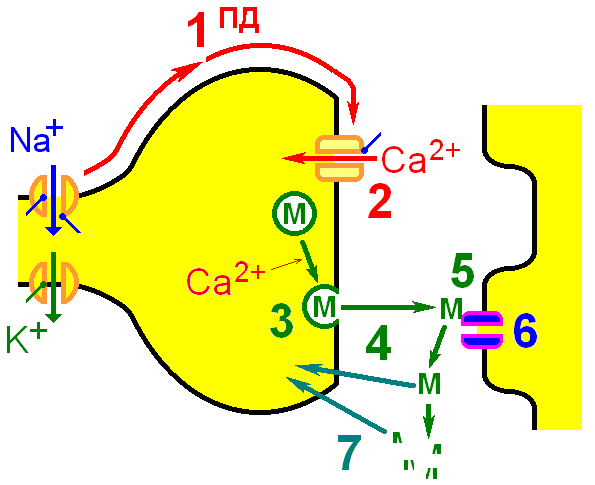
В 1953 году Бернард Катц с сотрудниками, исследуя нервно-мышечный синапс лягушки (медиатор - ацетилхолин), с помощью микроэлектродов зарегистрировали новый класс постсинаптических потенциалов
Эти сигналы возникали случайно, в покое, имели очень маленькую амплитуду и были названы миниатюрными потенциалами концевой пластинки (МПКП). Далее оказалось, что постсинаптические потенциалы, вызванные раздражением двигательного нерва (потенциалы концевой пластинки - ПКП), от раздражения к раздражению варьируют по амплитуде, причем эти колебания кратны амплитуде МПКП. Было предположено, что медиатор в синапсе освобождается в виде мультимолекулярных порций - квантов.
В покое случайное освобождение из нервного окончания отдельных порций вызывает появление на постсинаптической мембране МПКП, а в ответ на раздражение происходит синхронное освобождение нескольких десятков или сот квантов и возникает ПКП. Электрофизиологическое определение показало, что квант медиатора состоит из 1000-10 000 молекул ацетилхолина. В дальнейшем квантовая гипотеза освобождения медиатора получила подтверждение на самых разнообразных объектах и в настоящее время считается общепризнанной.
Использование метода электронной микроскопии для изучения ультраструктуры синапса позволило в 1954 году Де Робертсу и Беннету выявить в цитоплазме двигательного нервного окончания большое количество синаптических везикул диаметром около 50 нм. Поскольку везикулы имели одинаковые размеры и концентрировались у пресинаптической мембраны, было предположено, что квант медиатора находится в синаптической везикуле, а освобождение медиатора происходит путем выделения содержимого везикулы в синаптическую щель путем экзоцитоза.
Так была сформулирована везикулярная гипотеза освобождения медиатора в синапсе. В дальнейшем синаптические везикулы были обнаружены во всех химических синапсах нервной системы.
К основным постулатам везикулярной гипотезы необходимо отнести следующие: 1) медиатор в нервном окончании концентрируется в синаптических везикулах, 2) везикулярный медиатор освобождается путем слияния мембраны везикулы с пресинаптической мембраной (экзоцитоз).
Типы везикул. Как показали электронно-микроскопические исследования, самые разнообразные нервные окончания имеют два типа секреторных везикул: синаптические везикулы (мелкие везикулы) и секреторные гранулы (так называемые крупные или электронно-плотные везикулы). Мелкие синаптические везикулы однородны по размерам и имеют малый диаметр (около 50 нм). Эти везикулы содержат классические медиаторы. Крупные, электронно-плотные везикулы имеют большой диаметр (около 100 нм), они неоднородны по размерам и содержат электронно-плотные гранулы, представляющие собой крупномолекулярные медиаторы - пептиды и белки.