

3.5.Вторичные мессенджеры (внутриклеточные посредники) гормонов. Циклический амф, его образование и распад. Аденилатциклаза и фосфодиэстераза. Действие ц-амф на протеинкиназы (адреналин, глюкагон).
Гормоны
влияют на активность ферментов, выступая
в роли аллостерических эффекторов. Это
так называемый механизм срочной
регуляции. Его осуществляют гормоны,
взаимодействуя с мембранными рецепторами
пептидные гормоны, кальцитонин, глюкагон,
паратгормон, вазопрессин и адреналин,
а также гормоны местного действия –
цитокины– низкомолекулярные белки,
синтезирующиеся в процессе иммунного
ответа, являющиеся медиаторами иммунной
и воспалительной реакций, факторы роста
и дифференциации клеток.
Эйкозаноиды-
местные гормоны, синтезирующиеся из
арахидоновой кислоты- избыточная
продукция при многих заболеваниях.
Гормон,
взаимодействуя с рецептором, образует
комплекс, который приводит к активизации
G-белка, что в свою очередь активизирует
АЦ (в молекуле которой много аллостерических
центров). Она катализирует расщепление
АТФ с образованием цАМФ. Он активирует
неактивную протеинкиназу, а та- неактивную
фосфорилазу «в» в активную фосфорилазу
«а», способствующую распаду гликогена
до глюкозы, уровень которой в крови
повышается.
Другая система, генерирующая
цГМФ, сопряжена с ГЦ. Молекулы цГМФ могут
активировать протеинкиназу G, участвующую
в фосфорилировании других белков в
клетке. Например, ФДЭ активируется в
результате фосфорилированияцГМФ-
зависимой протеинкиназой G.
Синтез
цАМФ и цГМФ четко регулируется так
называемыми устраняющими механизмами:
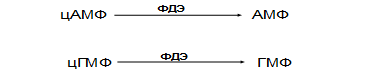 Образующиеся
свободные АМФ и ГМФ не способны
активировать ферменты.
Такие гормоны,
как вазопрессин и адреналин, образуя
комплекс с соответствующим рецептором
через активацию соответствующего
G-белка, активируют фосфолипазу «С», в
результате чего в клетке появляются
вторичные посредники ДАГ, ИФ3.
Молекула ИФ3 стимулирует
высвобождение Са2+ из ЭР
(эндоплазматический ретикулум). Са2+
связывается с белком кальмодулином.
Этот комплекс активирует Са2+-
кальмодулинзависимуюпротеинкиназу.
Ионы Са2+ и ДАГ активируют
протеинкиназу «С».
Образующиеся
свободные АМФ и ГМФ не способны
активировать ферменты.
Такие гормоны,
как вазопрессин и адреналин, образуя
комплекс с соответствующим рецептором
через активацию соответствующего
G-белка, активируют фосфолипазу «С», в
результате чего в клетке появляются
вторичные посредники ДАГ, ИФ3.
Молекула ИФ3 стимулирует
высвобождение Са2+ из ЭР
(эндоплазматический ретикулум). Са2+
связывается с белком кальмодулином.
Этот комплекс активирует Са2+-
кальмодулинзависимуюпротеинкиназу.
Ионы Са2+ и ДАГ активируют
протеинкиназу «С».
4.5.Кальций как вторичный мессенджер гормонов. Кальмодулин. Образование и действие инозитолтрифосфата (иф3)и диацилглицерола (даг) как внутриклеточных сигнальных компонентов действия гормонов
вазопрессин и адреналин, образуя комплекс с соответствующим рецептором через активацию соответствующего G-белка, активируют фосфолипазу «С», в результате чего в клетке появляются вторичные посредники ДАГ, ИФ3. Молекула ИФ3 стимулирует высвобождение Са2+ из ЭР (эндоплазматический ретикулум). Са2+ связывается с белком кальмодулином. Этот комплекс активирует Са2+- кальмодулинзависимуюпротеинкиназу. Ионы Са2+ и ДАГ активируют протеинкиназу «С».
5.5. Гормоны коры надпочечников. Химическое строение. Изменение метаболизма при гипер-, гипокортицизме (болезнь Аддисона, болезнь Иценко-Кушинга).
У 2-х месячного эмбриона масса надпочечников равна массе почки и представлена в основном корой, продуцирующей андрогены (фетальная кора). Мозговое вещество еще не выявляется. После рождения и до года жизни идет уменьшение массы надпочечников до 4-3,5 г. После 1 года жизни вновь начинается увеличение массы надпочечников и к 10-15 годам их масса достигает 8-9 г. Фетальная кора полностью исчезает к 3-4 годам.
Функциональная активность коры надпочечников в различные периоды жизни неодинакова. С мочой в первые два – три дня у новорождённого выводятся в основном метаболиты материнских гормонов. К 4-му дню жизни ребенка происходит снижение экскреции и продукции стероидов. В это время возможно проявление клинических признаков недостаточности надпочечников. Но уже к 10-му дню жизни происходит активация синтеза гормонов коры надпочечников и улучшение общего состояния ребенка.
Общим предшественником всех кортикостероидов служит холестерин. При его синтезе образуется более 40 метаболитов, различающихся по структуре и биологической активности. Основными кортикостероидами, обладающими выраженной гормональной активностью, являются кортизол – глюкокортикоид, альдостерон - минералокортикоид и андрогены. Стероидные гормоны транспортируются кровью в комплексе со специфическими транспортными белками. Скорость синтеза и секреции кортизола регулируется гипоталамо-гипофизарной системой по механизму обратной связи.
Глюкокортикоиды повышают уровень глюкозы в крови, по механизму хронической регуляции способствуют синтезу ферментов глюконеогенеза. Минералокортикоиды – задерживают ионы Na+, Cl¯ и воду в организме ( увеличивая обратное всасывание) и способствуют выведению из организма ионов К+.
Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов. Эти продукты метаболизма выделяются преимущественно в виде коньюгатов с глюкуроновой и серной кислотами.
У мужчин 2/3 кетостероидов образуется за счет кортикостероидов и 1/3 – за счет тестостерона (всего 12-17 мг/сут). У женщин 17-кетостероиды образуются преимущественно засчет кортикостероидов (12-17 мг/сутки).
Гипофункция гормонов коры надпочечников (гипокортицизм) – болезнь Аддисона или бронзовая болезнь развивается в результате туберкулезного или аутоиммунного повреждения. Клинические проявления: гипогликемия, выделение из организма большого количества ионов Na+, Cl¯ и воды (поносы), обезвоживание, гипотония (понижение артериального давления), задержка в организме ионов К+. При этом наблюдается усиленная пигментация кожи. Это связано с тем, что по механизму обратной связи в гипофизе вырабатывается много АКТГ, который стимулирует выработку пигмента кожи меланина.
Гиперфункция гормонов коры надпочечников (гиперкортицизм) – болезнь Иценко-Кушинга или стероидный диабет, сопровождающийся гипергликемией, глюкозурией, в организме в большом количестве задерживаются ионы Na+, Cl¯ и воды. Это приводит к отекам (лунообразное лицо), к повышению артериального давления.
6.5. Гормоны поджелудочной железы. Инсулин,глюкагон, структура. Механизм их действия. Сахарный диабет. β-клетками поджелудочной железы вырабатывается гормон инсулин из предшественника проинсулина. В свою очередь проинсулин образуется из препроинсулина. Превращение предшественников в инсулин происходит в процессе частичного протеолиза.
Молекула инсулина состоит из двух полипептидных цепей, соединеных между собой в двух точках дисульфидными мостиками. Цепь А состоит из 21 аминокислотного остатка, цепь В – из 30 аминокислотных остатков.
Гормон инсулин вырабатывается, когда уровень глюкозы в крови повышается (гипергликемия), при этом он понижает содержание глюкозы.
Инсулин увеличивает проницаемость клеточных мембран по отношению к глюкозе. Инсулин дисульфидными мостиками соединяется с SH-группами мембраны клетки и в результате из пластинчатой мембраны образуется глобулярная и глюкоза проходит внутрь клетки.
Инсулин активирует по типу аллостерического взаимодействия фермент гексокиназу, которая катализирует превращение глюкозы в глюкозо-6-фосфат.
гексокиназа в печени
Г
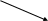 люкоза
глюкозо-6-фосфат
гликоген
люкоза
глюкозо-6-фосфат
гликоген
в тканях
источник энергии
Инсулин по механизму хронической регуляции является индуктором синтеза фермента гексокиназы и репрессором синтеза ферментов глюконеогенеза.
При недостаточном синтезе инсулина возникает инсулинзависимый диабет или диабет первого типа. Уровень глюкозы в крови увеличивается (гипергликемия), глюкоза появляется в моче (глюкозурия), повышается содержание в крови гликозилированного гемоглобина. В связи с этим у больных сахарным диабетом появляются следующие симптомы: полиурия (частое мочеиспускание), так как глюкоза повышает осмотическое давление крови; полидипсия (жажда, сухость во рту); полифагия (частое употребление пищи, голод), так как глюкоза не окисляется и возникает дефицит энергии.
Гормон глюкагон, состоящий из 29 аминокислотных остатков, секретируется α-клетками поджелудочной железы. Он образуется из предшественника – проглюкагона, который в процессе частичного протеолиза превращается в глюкагон. Глюкагон вырабатывается, когда уровень глюкозы в крови уменьшается (гипогликемия). Глюкагон повышает уровень глюкозы в крови за счет распада гликогена, активируя фермент фосфорилазу по механизму срочной регуляции (аденилатциклазная система). Кроме того, глюкагон стимулирует образование глюкозы из аминокислот путем индукции синтеза ферментов глюконеогенеза.