

8.4.. Иммобилизованные ферменты, применение в медицине. Изоферменты лактатдегидрогеназы. Значение определения изоферментов в диагностике заболеваний.
Практическое использование ферментов столкнулось с большими трудностями. Первое – это сложность и дороговизна получения достаточных количеств ферментов в чистом виде. Кроме того, ферменты быстро теряют свою активность под действием различных факторов (изменение кислотности среды, температуры, солевого состава и др.)
При использовании ферментов возникают осложнения, в первую очередь иммунологические. Также невозможно создать высокую местную концентрацию фермента при локальных поражениях, так как ферменты не обладают способностью к «направленному транспорту».
Группой под руководством отечественного ученого И. Березина впервые были созданы иммобилизованные ферменты.
Под иммобилизацией ферментов понимают их физическое (адсорбционное) или химическое (ковалентное) связывание с матрицей носителя, которая защищает фермент от инактивирующих воздействий, но в минимальной степени влияет на функционирование его активных центров. Материалом для изготовления носителя могут служить неорганические пористые стекла, силикагели, а также природные или синтетические полимеры.
Разработаны несколько видов иммобилизации.
Наиболее распространены реакции ацилирования, в которые могут вступать амино-окси- и некоторые другие группы белка, при этом чаще всего реакция протекает по аминогруппам лизиновых остатков.
Очень распространена реакция образования азаметиновой связи (оснований Шиффа) между альдегидными группами носителя и аминогруппами белка.
Относительно новый способ получения устойчивых терапевтических ферментов – иммобилизация на соединениях, характерных для самого организма или даже обладающих собственной биологической активностью, дополняющих или усиливающих действие связанного с ними фермента. Примером может служить фибринолизин, иммобилизованный на гепарине, урокиназа – на альбумине.
Наконец, растворимые препараты иммобилизованных ферментов медицинского назначения могут быть получены путем их межмолекулярного слияния. Например, слияние молекул галактозидазы обеспечивает стабилизацию фермента. Одновременно замедляется переваривание фермента и увеличивается время его нахождения в кровотоке.
Иммобилизованные ферменты имеют ряд преимуществ. Они обладают достаточно длительным сроком годности, у них снижена аллергичность и иммуногенность за счет частичной или полной блокады антигенных участков белка макромолекулой носителя, они слабо восприимчивы к действию естественных ингибиторов и проявляют терапевтическую активность в течение длительного времени. Носитель, с которым связан фермент, обеспечивает не только устойчивость, но и направленную доставку фермента предпочтительно в зону поражения, т.е. в определенный орган или ткань.
Область применения иммобилизованных ферментов очень широка. Они могут быть компонентами аналитических систем для клинического биохимического анализа, могут служить для модификации внутренних поверхностей, как различного рода протезов, так и медицинских аппаратов, могут являться компонентами перевязочных и дренирующих материалов, обеспечивающих ускорение заживления и очищения ран.
Если фермент служит для лечения местных поражений (опухолей, тромбов) и его присутствие в других органах нежелательно, то создаются биосовместимые и биоразлагаемые производные ферментов в виде микрочастиц, гранул, таблеток.
Некоторые иммобилизованные ферменты используются для наружного применения, при включении в состав различных мазей или кремов.
Изоферменты – это множественные формы одного и того же фермента. Изоферменты катализируют одну и ту же реакцию, но отличаются по аминокислотному составу и некоторым физико-химическим свойствам (молекулярной массе, электрофоретической подвижности и др.). Например, фермент лактатдегидрогеназа (ЛДГ) существует в пяти формах: ЛДГ1, ЛДГ2, ЛДГ3, ЛДГ4, ЛДГ5.
ЛДГ катализирует реакцию:
Г люкоза
люкоза
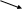 ЛДГ
4,5
ЛДГ
4,5


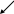 ПВК
молочная кислота +
2 АТФ
ПВК
молочная кислота +
2 АТФ
ЛДГ 1,2

СО2 + Н2О + 38 АТФ
Каждая ткань имеет определенный изоферментный спектр ЛДГ: в сердечной мышце – ЛДГ1,2; в печени, скелетных мышцах – ЛДГ4,5.
Какова целесообразность синтеза фермента в нескольких молекулярных формах? В тканях с преимущественно аэробным обменом веществ (сердечная мышца), преобладают формы ЛДГ1,2, которые обеспечивают ткани большим количеством энергии. В тканях с преимущественно анаэробным обменом веществ (печень, скелетные мышцы) преобладают ЛДГ4,5, что приводит к образованию молочной кислоты и двух молекул АТФ.
Определение уровня активности изоферментов в сыворотке крови имеет важное значение в диагностике заболеваний. например, повышение активности ЛДГ1,2 наблюдается при инфаркте миокарда; ЛДГ4,5 – при заболеваниях печени (гепатит, цирроз).
При заболеваниях, сопровождающихся некрозом, маркерные (органоспецифичные) ферменты из повреждённых клеток в большом количестве поступают в кровь, и уровень их активности увеличивается, возникает гиперферментемия. Определение уровня активности маркерных ферментов в сыворотке крови имеет клиническое значение в диагностике и прогнозе ряда заболеваний.
1.5. Гормоны. Классификация, биологическая роль. Центральная регуляция эндокринной системы. Нервно-рефлекторный и эндокринный пути действия центров гипоталамуса на эндокринный аппарат. Либерины, статины. Строение, биологическая роль.
Гормоны – биологически активные вещества, образуемые железами внутренней секреции в следовых количествах, оказывающие регулирующее влияние на обмен веществ и физиологические функции.
Синтез и секреция гормонов стимулируются внешними и внутренними сигналами, поступающими в различные отделы ЦНС.
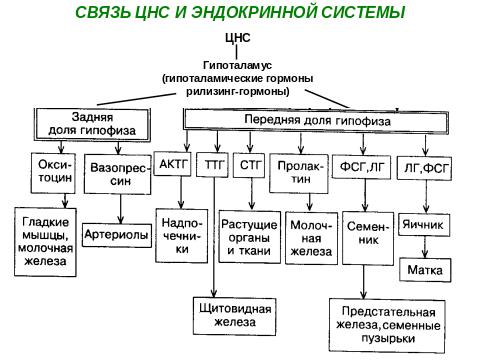
Эти сигналы по нейронам поступают в гипоталамус, где стимулируют синтез пептидных рилизинг-гормонов (от англ. release - освобождать).
7 либеринов активируют выработку гипофизарных гормонов и 3 статина, которые тормозят.
По химической природе это пептиды (состоят из 3,10,14 аминокислотных остатков).
Тиролиберин- трипептид (глутаминовая кислота, гистидин, пролин). (ТТГ и пролактин).

Соматостатин - 14 аминокислотных остатка- угнетает выработку СТГ гипофиза.
Тропные гормоны передней доли гипофиза стимулируют образование и секрецию гормонов периферических эндокринных желез, которые поступают в кровь и накапливаются в определенных органах-мишенях.
АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (минерало- и глюкокортикоидов).
ТТГ- синтез и секрецию гормонов щитовидной железы;
СТГ – на растущие органы и ткани и секрецию и синтез гормонов поджелудочной железы;
Пролактин – на секрецию молочных желез;
ФСГ, ЛГ – на секрецию семенников и яичников
Поддержание уровня гормонов в организме обеспечивает механизм отрицательной обратной связи. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус; синтез и секреция тропных гормонов угнетается гормонами периферических желез.
Такие петли обратной связи действуют в системах регуляции гормонов надпочечников, щитовидной железы, половых желез.
Но не все эндокринные железы регулируются подобным образом. Например, гормоны задней доли гипофиза (вазопрессин и окситоцин) синтезируются в гипоталамусе в виде предшественников и хранятся в гранулах терминальных аксонов нейрогипофиза.
Секреция инсулина и глюкагона напрямую зависит от уровня глюкозы.