

Отличия высших растений от низших.
Высшие растения отличаются от низших по многим признакам, однако главное их отличие в том, что они по преимуществу - обитатели суши, устроены несравненно более сложно и более разнообразны, чем низшие растения. В отличие от низших растений, высшие растения представляют собой сложно дифференцированные многоклеточные организмы, приспособленные к жизни в наземной среде (за исключением немногочисленных и явно вторичных водных форм), с правильным чередованием двух поколений: полового (гаметофит) и бесполого (спорофит). На спорофитах высшисх растений развиваются многоклеточные спорангии, в которых образуются неподвижные споры, на гаметофитах — многоклеточные половые органы (гаметангии), однако у некоторых голосеменных и у всех цветковых растений гаметангии в процессе эволюции исчезли.
Спорангии одинаковые или чаще дифференцированы на 2 типа (микроспорангии и мегаспорангии), гаметангии — всегда двух типов — мужские (антеридии) и женские (архегонии). Как антеридии, так и архегонии защищены многоклеточными стенками. Женская гамета (яйцеклетка) всегда одна, неподвижна. Зигота у высших растений развивается в многоклеточном зародыше, который проходит первые стадии развития внутри женского гаметофита. Спорофит у высших растений, как правило, расчленён на 3 основных органа — лист, стебель и корень. В большинстве случаев у высших растений развивается специальная проводящая ткань — ксилема и флоэма, у некоторых групп редуцировавшаяся. Имеется эпидерма с кутикулой и типичными устьицами. Хлорофилл у высших растений не содержит дополнительных пигментов, окраска фотосинтезирующих частей зелёная. Еще более показательны различия вклада высших и низших растений в продуктивность и биомассу биосферы. Низшие растения производят примерно 1/3 первичной биологической продукции, но ввиду того, что длительность жизни большинства из них невелика, биомасса низших растений составляет доли процента от общей массы продуцентов биосферы. Впрочем, разделение на высшие и низшие только по среде обитания будет не совсем верным. Часть высших растений ведет водный образ жизни, они называются гидрофитами. Среди гидрофитов много обитателей пресноводных водоемов (Nymphaeaceae, Lemnaceae, Potamogetonaceae и др.). В то же время, многие водоросли ведут наземный образ жизни и заселяют поверхность почвы и даже скальных пород. Однако в обоих случаях водная и наземная среда вторичны, то есть гигрофиты - это высшие растения, которые перешли от сухопутного к водному образу жизни, а наземные водоросли - водные растения, которые поселились на суше. Все водоросли могут нормально функционировать только в периоды, когда субстрат их обитания обильно увлажнен. В другое время они находятся в диапаузе.
Типы стели. Эволюция стели.
Стела занимает центральную часть стебля, состоит из проводящих тканей, сердцевины (иногда она разрушается), перицикла (если он имеется) и тех постоянных тканей, которые из него возникают. Кнаружи от перицикла (наружной границы стелы) располагается первичная кора.
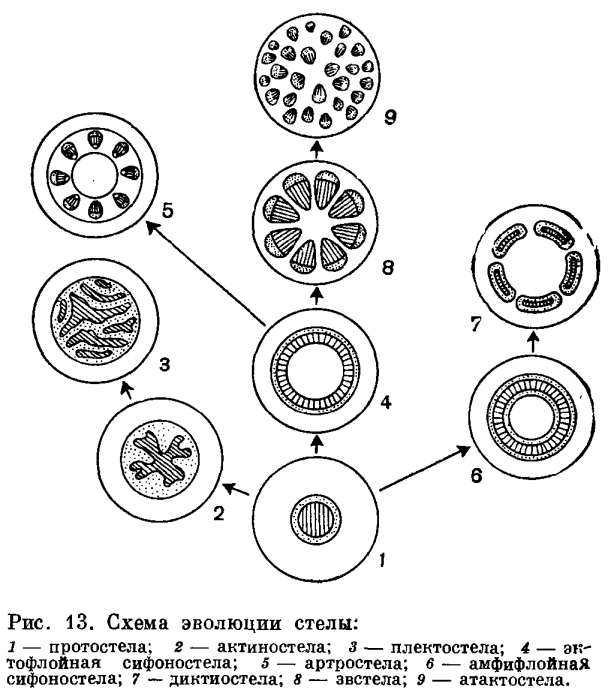
Протостела - наиболее примитивный тип стелы, в которой проводящая ткань образует сплошную массу и центральное ядро, состоящее из ксилемы, полностью окружено слоем флоэмы. Протостела не только очень проста по структуре, но и представляет собой исходный тип, от которого в процессе эволюции произошли все другие типы стелы. Она была характерна для риниофитов и многих других вымерших форм. В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой. Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой. Она встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела, характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле. В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела, которая, как и сифоностела, характерна для многих папоротников.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой. Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела, характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой. Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Жизненная форма растений, биологическая форма, биоморфа — внешний облик растений, отражающий их приспособленность к условиям среды. Форма, в которой вегетативное тело растения находится в гармонии с внешней средой в течение всей жизни, от семени до отмирания.
Жизненная форма растения не остается постоянной, а может меняться по мере взросления и старения. Важнейшую роль в становлении жизненной формы играет внешняя среда. Но это, конечно, не означает, что жизненная форма любого растения бесконечно пластична и зависит только от непосредственно действующих в данный момент условий.
Каждый вид растений реагирует на внешние воздействия в рамках своих наследственно закрепленных возможностей. Земляника, например, не станет развесистым деревом даже в самой благоприятной для роста и ветвления обстановке. Подразумевается, что в сложившейся жизненной форме каждого вида проявляются черты наследственной, выработанной в процессе естественного отбора приспособленности к определенному комплексу внешних факторов.
Использовав и обобщив предложенные в разное время классификации, отечественный ботаник И.Г. Серебряков предложил называть жизненной формой своеобразный габитус определенных групп растений, возникающий в результате роста и развития в определенных условиях – как выражение приспособленности к этим условиям.
В основу своей классификации И.Г. Серебряков положил признак продолжительности жизни всего растения и его скелетных осей. Он выделил следующие жизненные формы растений:
А. Древесные растения
Деревья
Кустарники
Кустарнички
Б. Полудревесные растения
Полукустарники
Полукустарнички
В. Наземные травы
Поликарпические травы (многолетние травы, цветут много раз)
Монокарпические травы (живут несколько лет, цветут один раз и отмирают)
Г. Водные травы
Земноводные травы
Плавающие и подводные травы
Различие между деревьями, кустарниками, кустарничками, полукустарниками, полукустарничками и травянистыми растениями состоит, помимо разной степени одревеснения их стеблей, в продолжительности жизни и характере смены скелетных побегов в общей побеговой системе.
Жизненные формы растений можно классифицировать по самым разным морфологическим признакам. В основу своей классификации Раункиер положил один единственный, но экологически значимый признак, - высоту залегания почек возобновления над поверхностью субстрата. На его основе Раункиер выделил пять крупных групп жизненных форм растений.
Фанеровиты - почки расположены достаточно высоко над поверхностью и в лучшем случае защищены почечными чешуями. К этой группе относят деревья и кустарники.
Хамефиты - почки располагаются на незначительной высоте (20-30см) над поверхностью. В эту группу входят кустарнички, полукустарники, стелющиеся растения, растения-подушки.
Гемикриптифы - почки возобновления находятся на уровне субстрата (дерновинные, розеточные и др. формы травянистых растений).
Геофиты - почки возобновления расположены в почве или другом твердом субстрате на некоторой глубине (корневищные, клубневые, луковичные растения).
Гидрофиты - почки возобновления размещаются в воде.
Терофиты - особая группа. Это однолетние растения, вегетативные части которых отмирают к концу вегетационного сезона, а зимующие почки вообще не образуются.
Лист – боковая структурная часть побега, как правило, характеризующаяся билатеральной симметрией, плагиотропным ростом и уплощенной формой. Он нарастает интеркалярно. Лист выполняет три главнейшие функции: фотосинтеза, транспирации и газообмена.
Первые листовые органы семенных растений – семядоли зародыша. Следующие листья формируются входе онтогенеза первоначально в виде меристематических бугорков, возникающих на апексе побега. Позднее бугорки развиваются в выступы – листовые примордии, а эти последние – в листья.
Существуют два принципиально возможных способа возникновения плоских органов фотосинтеза (листьев) на цилиндрическом теломе, и, вероятно, оба они реализовывались в эволюции растений. Один из них состоит в прогрессирующем развитие энациев, т.е. выростов поверхности тела с параллельным их уплощением. В результате появились многочисленные, густо покрывающие стебель мелкие листья крайне простой формы. В ходе последующей эволюции листья могли приобретать большие абсолютные размеры, но почти всегда сохраняли простую форму. Обладающие такими листьями растения наз. микрофилльными независимо от абсолютного размера листьев. Это – мохообразные, плаунообразные.
Другой способ возникновение листьев связан с функциональной дифференциацией ветвей дихоподиально или моноподиально нараставших теломов и наиболее полно прослежен у теломных риниевых, тримерофитовых, примитивных папоротникообразных. При этом лидирующие ветви специализировались на размещении в пространстве более слабых ветвей, занимавших боковое положение и в основном осуществлявших функцию фотосинтеза. Преобразование таких ветвей в листья включало несколько процессов:
Преобразование потенциально неорганиенного роста в ограниченный при интенсификации ветвления.
Плантация, преобразование трехмерного ветвления в плоскостное, уплощение веточек.
Агрегация всех ответвлений плоскости.
Срастание ее ответвлений.
Эти способом возникали не очень многочисленные листья, сохранявшие некоторые признаки ветвей: длительный апикальный рост, сложное расчленение и относительно крупный размер – макрофильные листья. В ходе последующей эволюции листья у многих растений из них стали мелкими и значительно упростили свою форму.
Строение: Взрослый лист обычно расчленен на пластинку или несколько пластинок (у сложных листьев) и черешок – узкую стеблевидную его часть, соединяющую пластинку и узел побега. Самая нижняя часть листа, сочлененная со стеблем, наз. основанием листа. Часто при основании заметны разного размера и формы парные боковые выросты – прилистники. Пластинка – главнейшая часть листа, осуществляющая его основные функции. Редуцируется пластинка весьма редко. И тогда ее функции принимают либо филлодий (расширенный листовидный черешок), либо крупные листовые прилистники. Нередко черешок не развивается, и тогда лист – сидячий. С черешком – черешковым.
У части видов растений хорошо выражена гетерофилия. Во гетерофилиею или ризнолистистю понимают разнообразие формы листьев, которые имеют примерно одинаковый возраст и расположены на одном и том же побеге. Чаще гетерофилия связана с экологическими условиями существования. Достаточно четко гетерофилия выражена в водной растения - стрелолиста. У него формируется три типа хорошо отличных листьев: подводные, в виде узких лентовидных пластинок; плавающие, с округлой пластинкой; воздушные, со стреловидным пластинкой. Гетерофилия также хорошо выражена в листьев шелковицы. В нее можно наблюдать листья примерно одинакового возраста как с целостной листовой пластинкой, так с пластинкой с разной степенью расчленения. Достаточно хорошо гетерофилия выражена в австралийских филодийних акаций, которые во влажных условиях имеют двичиперистоскладни листья, а в засушливые периоды в них развиваются филоиды с редуцированной листовой пластинкой и сплющенным черенком. Частным случаем гетерофилии является анизофилия, при которой своей морфологией отличаются листья, находящиеся в одном узле (часто такое явление характерно для плагиотропних побегов).
Листовая серия: Листья на одном побеге никогда не бывают одинаковыми. Сильнее всего различия между самыми первыми листьями семенных растений, или семядолями, развивающимися в основании главного побега и последующими его листьями, которые составляют листовую серию главного побега. Семядоли отличаются от листьев листовой серии особенностями возникновения. Листья одной листовой серии также неодинаковы. У одних видов все различия сводятся лишь к большей величине и несколько более сильному расчленению средних листьев, причем изменения в пределах листовой серии постепенны. У других растений на протяжении листовой серии наблюдается резкое изменение в строении и размерах листьев, что позволяет выделить 3 их формации. Листья низовой формации, или катафиллы, чешуевидные и состоят из разросшегося листового основания, а пластинка и прилистники отсутствуют или представлены маленькими рудиментами на верхушке катафилла. Листья срединной формации наиболее крупные и сложно устроенные, обеспечивающие фотосинтетическую функцию растения. В области соцветия(не на всех побегах) развиты гипсофиллы, мелкие слаборасчлененные листья верховой формации. Часто они представлены, как и катафиллы, Чешуевидными разросшимися основаниями с редуцированными прилистниками и пластинками.
Метаморфозы побега: диапазон метаморфозов побега очень широк: от небольшого уклонения от типичного строения до сильно измененных форм, в которых лишь с большим трудом удается распознать побег. Побеги листовых суккулентов мало отличаются от обычных побегов. Их отличия заключаются в толщине листьев, имеющих форму цилиндра, полуцилиндра или трехгранника (хавортия извилистая). У стеблевых суккулентов большую толщину имеет стебель, а листья имеют ординарное строение (молочай буйволова), изменены или редуцированы до колючих прилистников (молочай латексоносного) или полностью (астрофитум многоточеченый). Видоизмененные органы суккулентов запасают воду: у листовых – листья, у стеблевых – стебли. Функцию запасания ассимилянтов выполняют разнообразные метаморфизованные побеги. Кочан – гигантская покоящаяся открытая почка с утолщенной стеблевой частью (кочерыгой) и десятками не развернувшихся листьев. Запасные вещества депонированы и в кочерыге, и в листьях. Корневище – долговечный побег, в котором запасает ассимилянты стебель, листья – катафиллы, недолговечны. По длине корневища развиваются многочисленные узловые придаточные корни. Главный корень почти всегда не сохраняется. По мере роста корневища его базальная часть постепенно отмирает, вследствие чего корневищное растение медленно перемещается на другое место. 2 типа корневища: 1- удлиненные или ползучие. Имеют удлиненные и довольно тонкие междоузлия. Растут плагиотропно, быстро занимая большие площади субстрата (ползучее корневище осоки острой). 2-укороченные. Имеют укороченные междоузлия, могут расти плагиотропно, ортотропно или косо в зависимости от вида растения (купена многоцветковая). Корневища могут быть гипогеогенными – возникают из подземной почки, с самого начала существуют подземно (вороньий глаз четырехлистный); и эпигеогенными – формируются надземно, постепенно погружаясь в почву(у земляники корни втягивают первоначально ортотропный побег в почву). Каудекс – похож на укороченное ортотропное корневище, но при нем сохраняется мощный главный корень (люпин многолетний). Корневище отмирает от основания к верхушке, а каудекс отмирает из центра к периферии: в центре его образуется полость. Стеблевой клубень – терминальная часть закончившего продольный рост видоизмененного элементарного побега. Он всегда имеет укороченные очень толстые одно (одночленный клубень) или несколько (многочленный клубень) междоузлий и слабо развитые катафиллы, едва заметные у зрелого клубня. Придаточные корни обычно не развиваются. Недолговечны, переживают сезон покоя и отмирают только в следующем вегетационном сезоне. Гипокотильный клубень представляет собой шарообразно разросшийся гипокотиль, иногда с укороченным основание главного побега. Туберидий очень похож на клубень, но развивается из базальной части надземного побега, включающих одно или несколько утолщающихся междоузлий, запасающих воду и ассимилянты (многочленный туберидий лендробиума).
Листья в зависимости от вида растения опадают, высыхают или сохраняются в виде пленчатого покрова. Клубнелуковица – подземный укороченный метаморфизованный побег с толстым стеблем, запасающим ассимилянты, придаточными корнями, отрастающими с нижней стороны клубнелуковицы, и сохраняющимися засохшими основаниями листьев совокупности составляющими защитный покров (шафран весенний). Запасные вещества в луковице запасаются в чешуях листовой природы, а стебель преобразован в донце: он очень сильно укорочен, имеет форму диска и снизу несет придаточные корни. Запасающие чешуи мясистые и соответствуют либо целому катафиллу, либо только разросшемуся основанию зеленого ассимилирующего листа. Наружные чешуи луковицы последовательно отмирают и засыхают. Кладодии - видоизменённый побег растений с уплощённым длительно растущим стеблем, который выполняет функции листа. Настоящие листья на кладодии редуцированы, превращены в колючки или рано опадают. Происхождение кладодия из побега доказывается положением его в пазухе листа (обычно чешуевидного), а также образованием на нём цветков, которые никогда не возникают на настоящих листьях. Кладодии присущи преимущественно растениям, произрастающим на засушливых территориях (некоторые виды кактусов, спаржи, смилакса), и рассматриваются как приспособления для уменьшения испарения.
Кладодии, которые рано прекращают свой рост, называются филлокладиями. Филлокладии встречаются у растений различных семейств, распространённых главным образом в засушливых местностях. Полагают, что филлокладии, как и филлодии, служат приспособлением к уменьшению транспирации (иглица). Усик – жгутовидный ветвистый или неразветвленный орган, лишенный листьев и метамерного строения. Обладает длительным, ограниченным органом. У некоторых видов апекс усика преобразуется в дисковидную присоску. Колючки – это побеги с ограниченным продольным ростом, очень твердым одревесневающим стеблем и острой верхушкой. Колючки вначале олиствененны, несут пазушные почки или лишены листьев с самого начала.
Метаморфозы листа. Метаморфизировать может не весь лист, а только его часть. Мешковидные, или асцидиатные, листья возникают на растущих ветвях деревьев для улавливания и сохранения воды. Накопившуюся воду растение поглощает придаточным узловым корнем, врастающим в полость листа. Чаще всего эти листья служат ловчими органами. Насекомые не могут выбраться из-за своеобразно завернутого края листа и текстуры внутренней поверхности, которая допускает движение только от устья вглубь ловушки. Ловчие пузырьки пузырьчатки представляющие собой асцидиатные конечные доли многократно раздельных листьев, устроены наиболее сложно. Они активно улавливают плавающую в воде добычу. Имеют подвижный клапан, плотно замыкающий устье. Снаружи вокруг устья растопырены 4 антенны. Филлодии – уплощенные черешки, функционально замещающие пластинку простого листа или листочки сложного листа, которые развиты слабо или вообще отсутствуют. Им свойственно параллельное или дуговое жилковани, притом что жилкование пластинки – сетчатое. Листовые усики и колючки могут соответствовать всему листу или только его отдельным частям (некоторым листочкам и рахису сложного перистого листа, как усики гороха, или прилистникам, как колючки белой акации). Шипы представляют собой не метаморфизированные органы, а выросты поверхности стебля и листа (роза).
Метаморфозы корня. Особенность метаморфозов корней состоит в том, что очень многие из них отражают не изменения главных функций корня, а изменение условий их выполнения. Микориза – наиболее распространенный метаморфоз корня. Это симбиоз корня и сросшихся с ним гиф грибов, из которых растение получает воду с растворенными в ней минеральными веществами. Эндомикориза – гифы проникают в глубь корня (травы). Эктомикориза – гифы густо оплетают корень снаружи (дуб). Сам корень теряет способность к неограниченному росту, изменяется текстура его поверхости и внутреннее строение. Клубеньки - небольшие утолщения на корнях многих растений (в первую очередь бобовых), в которых находятся симбиотические азотфиксирующие бактерии. У бобовых растений это ризобии (корни картофеля). Коралловидные корни – симбиоз семенных растений с азотфиксирующими организмами. Слабо растут в длину, но обильно ветвятся, боковые корни остаются частично сросшимися с материнским корнем друг кс другом. Саговниковые вступают в симбиоз с цианеями, фиксирующими атмосферный азот. Дыхательные корни или пневматофоры снабжают кислородом корневую систему, находящуюся в аноксической почве или иле. Пневматофоры – это утолщенные отрицательо геотропичные корни с ограниченным ростом, выставляющиеся над субстратом и пронизанные огромным числом полостей, по которым кислород диффундирует к скелетным и сосущим корням (кипарис болотный). Ходульные корни – придаточные корни на надземных побегах, которые растут вниз, не ветвясь и не образуя корневых волосков, пока не достигнут субстрата, где обильно ветвятся (фикус каучуконский). Досковидные корниотходят боковые (редко – придаточные) корни некоторых тропических деревьев, отходящие от основания ствола, имеющие уплощенную по вертикали форму и служащие дополнительной опорой дереву. Корни-прицепки развиваются на узлах побегов немногих видов лиан и закрепляются на опоре. Быстро прекращают рост, не образуют корневых волосков, но образуют боковые корни. Котрактильные корни развиваются на корневищах и луковицах, заглубляют их в грунт. Они интенсивно растут в длину не ветвясь, а затем сильно замедляют продольный рост и обильно разветвляются на конце (лилия кудрявая). Запасающие корни сильно утолщены. Это боковые или придаточные корни – корневые шишки, или корневые клубни, либо главный корень (корнеплод), в формировании которого участвуют утолщенные гипокотиль и основание главного побега. Корневые колючки защищают растения от фитофагов. У некоторых эпифитных растений и пальм. Ассимилирующие корни. Бывают плоскими, неограниченно нарастающими и умеренно ветвящимися; и располагающимися в толще воды на узлах побега. Корневых волосков не образуют.
Почка. Почка - это зачаточный побег. Она состоит из укороченного стебля с тесно сближенными зачаточными листьями. На верхушке стебля находится конус нарастания, состоящий из образовательной ткани. За счет деления клеток конуса нарастания происходит рост стебля в длину, образование листьев и наружных почек. Снаружи почка защищена почечными чешуями, которые представляют собой видоизмененные нижние листья побега. По расположению на побеге почки бывают верхушечными и боковыми.
Верхушечная почка - это почка, находящаяся на верхушке побега, остальные почки - боковые. Их делят на пазушные и придаточные.
Пазушные почки регулярно возникают в пазухах молодых листовых зачатков вблизи верхушки материнского побега. Их расположение точно соответствует листорасположения. Поэтому зимой по почкам можно определить расположение листьев.
Боковые почки, которые развиваются вне пазухи на междоузлиях, корнях и листьях, называются придаточными. Они часто обеспечивают вегетативное размножение растений. Придаточные почки на листьях сразу развиваются в маленькие растеньица с придаточными корнями, которые отпадают от листа материнского растения и вырастают в новые особи. Эти почки называются выводковыми (бриофиллиум, росянки). Они могут возникать в пазухе листа и видоизменяться в луковички (лилия тигровая) и клубеньки (горец живородящий). Почки по строению не одинаковы. У большинства растений они закрытые (защищенные), т.к. снаружи имеют почечные чешуи, склеенные смолой (у хвойных), другими клейкими веществами (тополь), некоторые часто опущены. Есть растения с открытыми (незащищенными, голыми) почками. Они лишены почечных чешуек (калина, крушина).
По внутреннему строению различают следующие типы почек:
1) вегетативные - состоят из зачаточного стебля, чешуек, зачаточных листьев и конуса нарастания; 2) генеративные - цветочные, состоят из зачаточного стебля, чешуй и зачатков цветка или соцветия (бузина красная); 3) смешанные - состоят из зачаточного стебля, чешуек, зачаточных листьев и зачатков цветка или соцветия (яблоня, спирея). Генеративные и смешанные почки крупнее и более округлые, чем вегетативные. Почки, которые пребывают в состоянии покоя (осень - зима), а затем разворачиваются и дают новые побеги, называются зимующими или почками возобновления. За их счет происходит нарастание побегов.
Спящие почки - они находятся в течении ряда лет в состоянии покоя. Стимулом для их пробуждения является повреждение ствола.