

13. Главный комплекс гистосовместимости (гкгс)
Большая и важная группа генов, получивших в своё время не совсем благозвучное определение на основании того, что белки, кодируемые ими, были впервые выявлены при отторжении трансплантата, т.е. при тканевой несовместимости. Ныне установлено, что истинная роль этих белков — служить маркёрами идентичности на поверхности различных клеток, с которыми Т-лимфоциты взаимодействуют через собственные рецепторы в иммунной реакции.
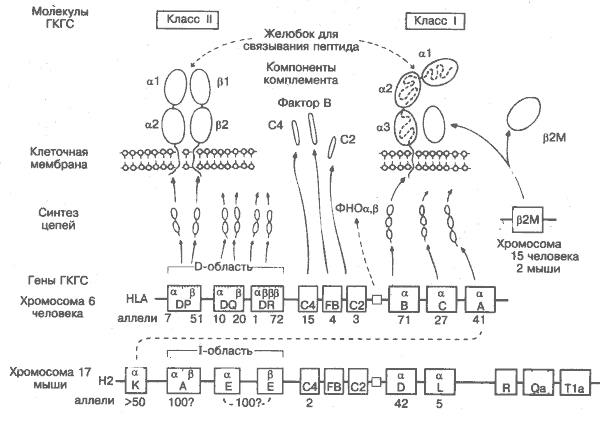
Исторически сложилось, что у мышей ГКГС известен как Н2, а у человека — как HLA (human leucocyte antigens). Изучение ГКГС у разных биологических видов пока не выявило существенных отличий по структуре и механизмам взаимодействия с Т-клетками. ГКГС состоит из группы генов класса I и класса II. На рисунке обозначения генов заключены в прямоугольники, под которыми указано число аллелей (альтернативных вариантов) для каждого локуса. ГКГС обладает огромным полиморфизмом, и только в пределах 1 хромосомы число возможных комбинаций её генов превышает 3-106. Молекулы ГКГС, кодируемые обеими хромосомами, создаются на основе 1 из почти 1013 комбинаций генов, что неимоверно осложняет пересадки органов, например почки (рис. 37).
Когда типирование HLA-системы получило широкое распространение, выяснилось, что от её состава зависит предрасположенность или, напротив, повышенная резистентность к многим заболеваниям. Было предложено несколько объяснений этой связи, но ни одно из них не является достаточно убедительным.
Н2 — мышиный ГКГС. расположенный на хромосоме 17. По крайней мере 20 других генов минорной гистосовместимости расположены на разных хромосомах и обозначены как H1. H3 и т.д., но только Н2-гены оказывают очень существенное влияние на отторжение трансплантата. Установлено также, что лишь они кодирует молекулы для нормальных межклеточных взаимодействий.
К, D, L — гены Н2 класса I, кодирующие a-цепь (м. м. 44 000), которая в сочетании с b2-микроглобулином образует четырёхдоменные молекулы К, D или L или антигены. N-концевые участки a-цепей крайне вариабельны и, возможно, эволюционировали для взаимодействия с различными вирусами. Поскольку вирусом может быть заражена любая клетка, то молекулы ГКГС класса I присутствуют на поверхности практически всех клеток организма, за исключением эритроцитов у некоторых биологических видов (в том числе у человека). На рисунке под каждым локусом проставлено число известных аллелей. Фактически их гораздо больше.
R, Qa, T1a считаются частью семейства класса I, так как кодируют очень сходные молекулы, которые представлены только на некоторых лимфоцитах и не опосредуют отторжение трансплантата.
А, Е — гены Н2 класса II, объединённые в I-область. А и Е содержат разные гены для а- (м. м. 33 000) и (3- (м. м. 28 000) цепей четырёхдоменной молекулы. В отличие от молекул класса I, молекулы класса II присутствуют только на клетках, взаимодействующих с Т-лимфоцитами и регулирующих их. Вероятно, огромное разнообразие молекул класса II объясняется существованием большего, чем показано на рисунке, количества генов А и Е со многими аллелями.
HLA — ГКГС человека, расположенный на хромосоме 6. Аналогичен Н2, за исключением того, что гены класса I расположены вместе и существуют как минимум 3 гена класса II.
А, В, С — гены HLA класса I, из них А — гомолог гена К у мышей. Существуют также гены HLA Е, F, G и Н, но функции их продуктов пока неясны.
DP, DQ, DR — гены HLA класса II. Продукты DP гена весьма эффективно стимулируют пролиферацию Т-клеток и выявляются в смешанной реакции лимфоцитов, тогда как все другие антигены классов I и II определяются антисыворотками. В этой же области находятся и другие гены (DN, DO, DZ), продукты которых пока неизвестны. Почти всю вариабельность, необходимую для взаимодействия с различными антигенами, а затем и с Т-клеточным рецептором, в молекулу класса II вносит b-цепь.
Обычно активно захваченный клеткой и расщеплённый в лизосомах до пептидов антиген связывается с молекулой ГКГС класса II, а антиген, образованный внутри клетки (например, из вируса), связывается с молекулой ГКГС класса I. Более подробно о взаимодействии антигенных фрагментов и молекул ГКГС см. на рис.14 и 17.
С2, С4, FB — гены ГКГС класса III. Кодируют компоненты комплемента, участвующие в активации СЗ. Интересно, что, хотя все они существуют в нескольких аллельных формах, значимых последствий этого не обнаружено. В этой же области находятся гены, кодирующие фермент надпочечников 21-гидроксилазу, цитокины ФНОa и b. Молекула, кодируемая С4, способна присоединяться к эритроцитам и мимикрировать под антигены группы крови.
М —микроглобулин (м. м. 12 000). Кодируется не в ГКГС, но входит в состав всех молекул класса I, стабилизируя их на поверхности клеток. В сыворотке обнаруживается в растворимой форме. У мышей имеет 2 аллельные формы, но в общем это одна из наиболее известных консервативных молекул.
ЗАБОЛЕВАНИЯ, СВЯЗАННЫЕ С HLA
Самый характерный пример — нарколепсия, наблюдаемая только у носителей DR2 антигена: впрочем, причины этого неизвестны. Интересны также артропатии крестцово-подвздошного сустава (анкилозирующий спондилит, болезнь Рейтера и др.), при которых HLA B27 аллель наблюдается в 95% случаев, т.е. почти в 20 раз чаще, чем обычно. Достаточно убедительно доказана связь многих других заболеваний, в том числе почти всех аутоиммунных, с частными антигенами или группами антигенов HLA (чаще всего D-области). Вероятнее всего, это объясняется различием в способности разных молекул HLA представлять микробные полипептиды или «свои» антигены. Возможно, имеется связь с генами других областей, особенно класса III. Тенденция некоторых HLA аллелей (например, А1 и В8) не разделяться, а оставаться сцепленными, называется неравновесным сцеплением. Возможно, такие комбинации являются жизненно важными, так как с ними может быть связана повышенная устойчивость к заболеванию или, напротив, повышенная чувствительность к нему. Вся эта область предоставляет широчайший простор для исследований.
14. Т-КЛЕТОЧНЫЙ РЕЦЕПТОР

Многие годы в иммунологии существовала ситуация, когда было известно, что Т-лимфоциты имеют на поверхности рецептор для антигена, сходный с антителами на В-лимфоцитах, но было неясно, насколько идентичны эти молекулы. Вопрос был снят в 1983-1984 гг., когда применение моноклональных антител и ДНК-зондов позволило точно установить, что и на молекулярном, и на генетическом уровне Т-клеточный рецептор уникален.
Рецептор Т-лимфоцита имеет характерную для суперсемейства иммуноглобулинов структуру, а именно состоящие из доменов полипептидные цепи, соединённые дисульфидными связями. Молекула рецептора имеет 2 основные цепи (a, p или, в отдельных случаях, у, 5), состоящие из 2 доменов каждая. Кроме того, во взаимодействии Т-лимфоцита с молекулами ГКГС участвуют молекулы CD3, CD4 и CD8. В правой части рисунка представлен процесс распознавания Т-хелпером чужого антигена в ассоциации с молекулой ГКГС. Сведения об этом процессе постоянно пополняются новыми данными. Особенно эффективно в этом отношении тестирование последствий генетического удаления («нокаутирование» гена) индивидуальных компонентов различных молекул, показанных на рисунке.
Необычность рецепторных белков состоит в том, что кодирующие их гены расположены на хромосоме не рядом, а через некоторые промежутки и для совмещения соответствующих генов происходит вырезание лежащих между ними сегментов ДНК, а потом РНК. Этот процесс, известный как перестройка генов, происходит только в Т-лимфоцитах, тогда как во всех других клетках гены остаются в нефункциональном состоянии зародышевой линии. Перестройка генов наблюдается в индивидуальном Т-лимфоците, что обеспечивает ему уникальный рецептор и как следствие уникальную антигенраспознаюшую способность. Аналогичный процесс происходит с генами иммуноглобулинов В-клеток (рис. 15). На этом и следующем рисунке серым цветом выделены участки генов и белков, которые, как считается, произошли из примитивного V(вариабельного)-региона (рис. 12), хотя и проявляют разную степень вариабельности.
ТКР — Т-клеточный рецептор. Состоит из одной a- (м. м. 50 000) и одной b- (м. м. 45 000) цепи, каждая из которых имеет внешний (вариабельный) и внутренний (константный) домены, внутримембранный и цитоплазматический короткий участки. На ранней стадии эмбрионального развития, а также в некоторых органах (кишечник, кожа) Т-лимфоциты могут иметь альтернативные рецепторы с gd-цепями и распознавать иные антигены по сравнению с ар Т-лимфоцитами. На рис. 10 даны происходящие в тимусе процессы позитивной и негативной селекции, в результате чего развиваются индивидуальные Т-лимфоциты, распознающие собственные молекулы ГКГС вместе с чужеродным пептидом.
CD3 — молекулярный комплекс, состоящий из 3 цепей: g (м. м. 25 000), d (м. м. 20 000) и e (м. м. 20 000), который играет существенную роль во всех функциях Т-клеток. Т-клеточный рецептор, CD3 и сигнальные двухцепочечные молекулы (zz и z,h) вместе образуют комплекс, взаимодействующий с антигеном (молекула ГКГС плюс пептид). Это взаимодействие ведёт к активации клетки по крайней мере через два внутриклеточных процесса, задействующих тирозинкиназу и фосфолипазу С, что в конечном счёте приводит к пролиферации клетки и высвобождению цитокинов.
CD4 — одноцепочечная поверхностная молекула (м. м. 60 000) Т-хелпера, участвующая в его взаимодействии с молекулами ГКГС класса II. CD4 является рецептором, через который ВИЧ проникает в Т-клетку (рис. 40).
CD8 — поверхностная молекула большинства цитотоксических Т-лимфоцитов, участвующая во взаимодействии с молекулами ГКГС класса I. CD8 человека состоит из 2 одинаковых цепей, её мышиный аналог — из 2 разных (Ly 2/3).
Подчеркивая тесную связь с Т-клеточным рецептором, молекулы CD4 и CD8 иногда называют корецепторами.
CD2, CD28, LFA-1 — три из многих адгезивных молекул, поддерживающие контакт Т-лимфоцита с антигенпредставляющими клетками или В-лимфоцитами (рис. 12). Особую роль играет молекула CD28, распознающая молекулу В7 как существенный костимулятор Т-клеточной активации. Считается, что в отсутствие костимуляции Т-клетка приобретает состояние неотвечаемости, которое может быть важным при развитии аутореактивности.
Перестройка генов. Гены ТКР содержат до 100 V-генов и множество J-генов. Для формирования отдельной цепи необходимо, чтобы по одном гену из них соединялось с С-геном. Это осуществляется вырезанием лежащих между ними сегментов ДНК. Подобный же процесс в информационной РНК заканчивается созданием конечной VDJC РНК, кодирующей полипептидную цепь. С учётом всех возможных комбинаций a- и b-цепей общее число различающихся молекул ТКР в организме может достигать 1010. CD4- и CDS-гены хотя и имеют явное V-генное происхождение, не перестраиваются и кодируемые ими молекулы не обладают таким разнообразием.
Антиген. В случае, представленном на рисунке, антиген в виде короткого пептида распознаётся молекулой ГКГС класса II и Т-клеточным рецептором (рис. 17). Достаточно полное распознавание с вовлечением взаимодействия CD4—молекула ГКГС класса II приводит к изменению молекулы CD3, в результате чего возрастает уровень внутриклеточного кальция и, возможно, других ионов. Т-лимфоцит активируется для выполнения своих запрограммированных функций. В случае активации цитотоксического Т-лимфоцита молекула CD8 связывается с молекулой ГКГС класса I, присутствующей на клетке-мишени.
ИЛ-2 — интерлейкин-2. Лимфокин, необходимый для начальной пролиферации Т-клеток, также действует на В-клетки, макрофаги, эозинофилы и др. (рис. 23). Зависимость между природой активирующего сигнала и разновидностью синтезируемого цитокина пока не совсем ясна. Эксперименты с клонируемыми Т-клетками, особенно на мышах, дают основания для заключения, что некоторые CD4 Т-клетки (Т-хелперы типа 1) продуцируют главным образом ИФ-7 и другие цитокины, активирующие макрофаги, хотя клоны других CD4 Т-клеток (Т-хелперы типа 2) преимущественно вырабатывают факторы, помогающие в антителопродукции (ИЛ-4, ИЛ-5, ИЛ-6 и др.). В настоящее время такое разграничение функций CD4 Т-клеток наблюдается у человека.
Суперантигены Имеется одно исключение в очень высокой специфичности взаимодействия Т-клетка—пептид—ГКГС. Некоторые вирусы и стафилококковые энтеротоксины обладают способностью связываться с b-цепью ТКР и молекулой ГКГС класса II вне пептидсвязывающего участка. В результате активируется не единственный клон Т-лимфоцитов, а целое семейство клонов, что приводит к повышенной продукции цитокинов с возможными повреждающими эффектами.