

5.Гібридизація і гетерозис
Гібрид (від грец.hibrida) — помісь— це гетерозиготний організм, одержаний в результаті схрещування генетично різних батьківських форм (видів, порід, ліній тощо). У широкому розумінні — кожна гетерозигота є гібридом.
Термін гетерозис (від грец.heteroiosis — зміна, перетворення) у 1911 р. запропонував Г. Шелл для позначення прояву високої гибридної сили нащадків, одержаних від схрещування різнорідних батьківських форм.
У 1905 р. під впливом досліджень Йогансена Г. Шелл та (окремо) Е. Іст спробували визначити реакцію особин кукурудзи (облігатна перехреснозапильна рослина) на примусове самозапилення. Вони показали, що при розмноженні вихідних гетерозиготних рослин таким шляхом нащадки розщеплюються на депресовані лінії, більшість яких гине. Але значна частина нащадків виживає. З'ясувалось, що від схрещування де-пресованих особин різних ліній одержували гібридних нащадків, які характеризувались високою життєздатністю й плодючістю, суттєвим збільшенням сили росту, підвищеною стійкістю до несприятливих умов середовища тощо. Таке різке зростання гібридної сили в гібридних нащадків й дістало назву гетерозису.
Вивчення гетерозису дозволило виявити ряд закономірностей цього явища. Зокрема, було встановлено, що сплеск який проявився в першому гібридному поколінні, згасає в наступних поколіннях, а від схрещування між генетично різними лініями утворюються сукупності нащадків з різними рівнями гетерозису. Останнє означає, що не кожна комбінація партнерів при схрещуванні здатна відтворити нащадків з високим гетерозисом. З'ясувалось, що із сотень гібридних комбінацій лише поодинокі забезпечують надвисокий сплеск гетерозису.
Зазначимо, що механізми гетерозису все ще вивчені слабо. Існує кілька теорій гетерозису. Одна з них - теорія домінування, запропонована Г. Девенпортом (в 1908 р.) і розвинена Д. Джонсом (в 1917 р.) Автори цієї теорії вважають, що гетерозис обумовлений дією сприятливих домінантних факторів, які які накопичені у групах зчеплення гібрида і забезпечують гетерозиготам (з цими факторами) деяку перевагу над гомозиготами. Проте згідно з теорією домінування гібридна сила і гетерозиготність пов'язані між собою не прямо, а опосередковано. Тому вважаються, що гетерозис — це наслідок кумулятивної дії багатьох домінантних генів, здатних посилювати інтенсивність росту й розвитку організмів.
Теоретично домінування, можна досить легко створити інбредні лінії з накопиченими в їхніх групах зчеплення домінантними генами, але досліди показали, що в таких лініях гетерозис аж ніяк не проявляється. Це й спонукало вчених переглянути цю теорію.
За уявленнями Г. Шелла (1911), І. Іста (1936), Ф. Хела (1952), С. Емерсона (1952) та інших дослідників для пояснення механізмів гетерозису була створена теорія гетерозиготності. Згідно з цією теорією надвисока гібридна сила у гібридних нащадків обумовлюється перевагами гетерозиготного стану генів над гомозиготним.
Теорія гетерозиготності, на відміну від теорії домінування, визнає, що обидва алельні гени кожного локуса в гетерозиготному стані активно функціонують і беруть участь у формуванні ознаки. Крім того, визнається, що молекулярні механізми гетерозиготності нездатні закріплювати гетерозис, тимчасом теорія домінування припускає таку можливість.
Гіпотеза моногібридного гетерозису. Моногібридний гетерозис проявляється в тому разі, коли гетерозигота стає суттєво сильнішою у формуванні певної ознаки, а ніж домінантна чи рецесивна гомозигота, тобто АА <Аа> аа. Якщо таке трапляється, то відповідну форму гетерозису називають, наддомінуванням або моногібридним чи моногенним гетерозисом.
Наявність моногенного гетерозису вважається доведеною, якщо він проявився в одній з двох ізогенних ліній, котрі відрізняються між собою лише за одним геном. У практиці дослідної роботи трапляються випадки виникнення спонтанних мутацій в гаметах особин, що належать до складу інбредної, або чистої, лінії. Внаслідок цього серед потомків нового покоління з'являються особини, які відрізняються від генотипів своїх сородичів лише одним мутантним геном, котрий знаходиться в гетерозиготному стані. Саме в таких особин очікується прояв моногібридного гетерозису, і очікувані ситуації мають місце.
Наприклад, в одній з чистих ліній ячменю була виявлена безхлорофільна мутація а(albina 7) і мутація х (xanta 3). Це летальні мутації. Але в гетерозиготному стані кожна з них обумовлює зростання врожаю зерна ячменю на 6%, а в гетерозигот а/+ х/+ урожайність зерна зростає на 15%. Отже, в даному разі проявився моногенний гетерозис.
Норки з сапфіровим забарвленням хутра гомозиготні за a1a1pp — рецесивними генами. В тварин АІАІрр — генотипу формується платинове забарвлення хутра. З'ясувалось, що введений в генотиплінії сапфірових норок домінантний ген АІ обумовлює формування платинового забарвлення хутра. Разом з цим підвищується плодючість самок і життєздатність їхніх нащадків. Тобто проявляється моноген-ний гетерозис, який досить ефективно використовують у хутряному звірівництві за схемою:
Отже, виробничники одержують високожиттєздатну сукупність коштовних кольорових норок, завдяки чому суттєво зростає ефективність виробництва. Гетерозис широко розповсюджений у природі. Він позначається на формуванні і розвитку ознак найрізноманітніших видів організмів. Вивчаючи закономірності уcпадкування ознак гетерозису, людина ефективно використовує їх у практиці селекційної та виробничої діяльності

Рис. Кон'югація гомологічних хромосом у Drozophila azteca, гетерозиготних за інверсіями (по Ф. Добжанському і М. Соколову). Зліва і справа одинокі інверсії, в центрі — подвійня інверсія, яка заходить одна за одну. Цифрами позначені ділянки політенних хромосом відповідно їх кросоверної карти.
Іноді та чи інша комбінація схрещування спричиняється до аномального розвитку ознаки, що призводить до її надмірного прояву. Така надмірність робить відповідні організми конкурентно- неспроможними в природних умовах. Проте іноді ці ознаки можуть використовуватись людиною у виробничій діяльності. Наприклад, ознаки прояву махровості, розмірів, забарвлення та запаху квітів — у квітникарстві; розміри та форма плодів у садівництві та овочівництві. При розщепленні гібридних нащадків у злакових рослин нерідко відщеплюються окремі особини зі збільшеними розмірами вегетативних і репродуктивних органів, але вони виявляються конкурентно-неспроможними. Отже, все це дає підстави вважати, що надмірний гетерозис виникає внаслідок несприятливих для виживання комбінацій генетичного матеріалу.
У другій половині 30-х років проводилися широкі дослідження з вивчення генетичних закономірностей життєдіяльності популяцій різних видів дрозофіл (М.П. Дубініним, Г. Тиняковим та М. Соколовим у колишньому СРСР; А. Стер-тевантом і Ф.Добржанським у США). Було виявлене дуже широке розповсюдження інверсій в каріотипах різних видів дрозофіл. На цій підставі зроблено висновок, що інверсії підхоплюються природним добором.На рис.показана будова деяких інверсій, котрі проявляються на політенних хромосомах.
Ф. Добржанський з співробітниками в дослідженнях на мухах Огозорпііа рзеисІооЬзсига (1960 р.) встановили, що популяції цього виду дрозофіл відрізняються між собою типами інверсій. При цьому Ф. Добржанський звернув увагу на те, що деякі інверсії в гетерозиготному стані характеризуються проявом високої гібридної сили. З'ясувалось, що ряд інверсій, проходячи через мейоз у процесі гаметогенезу, стабільно зберігають і успадковують свою вихідну цілісніть. Пояснюється це тим, що локалізовані в таких інверсіях комбінації різних генів не перекомбіновуються в кожному наступному мейозі і уcпадковуються в поколіннях.
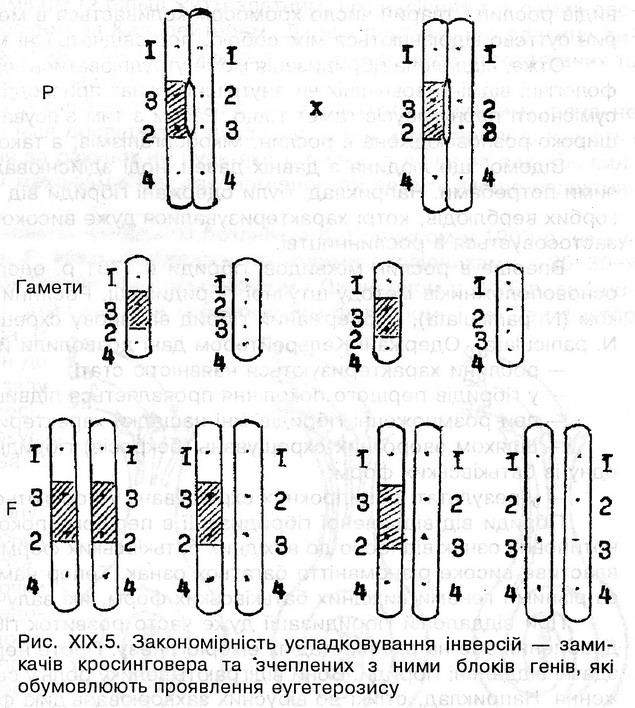
Механізмом стабільного успадкування гібридної сили в поколіннях є утворення мікроінверсій. які виконують функцію замикачів кросинговеру (рис. ХІХ.5). Якщо в утвореній мікроінверсії локалізований блок сприятливих для гетерозису генів, то цей гібридний стан успадковується в наступних поколіннях без роз щеплення (Добржанський назвав його еугетерозисом, тобто істинним гетерозисом). Таким чином, еугетерозис відрізняється від попередніх форм гетерозису тим, що спадкові структури, які його обумовлюють, успадковуються без розщеплення від покоління до покоління.