

10.04.2013 №8
Сестринские хроматидные обмены.
Между сестринскимихроматидами возникает реципропный обмен, сходный с кроссинговером, но не приводящий к появлению новых аллельных комбинаций. Сестринские ХО можно обнаружить с помощью спец методов окраски хромосом, при которых можно различить хроматиды. После такой окраски некот хромосомы выглядят мозаичными за счет многочисленных межхроматидных обменов - хромосомы типа Арлекин.
Частота СХО ниже, чем кроссинговер, и она изм-ся под действием мутагенов и при некоторых наследственных заболеваниях (синдром Блума: задержка роста, чувствительность кожи к УФ, предрасположенность к опухолям).
Соматический кроссинговер.
В некот случаях в сомат клетках происходит кроссинговер (митотич/соматич).
Доказательство: Штерн 1936, у мух, гетерозиготных по генам окраски тела иформе щетинок, наблюдались мозаичные пятна, кот предст собой группу клеток, фенотипически отличающихся от ост клеток и сосед мозаич пятна.
При наличии СК на сером теле с норм щетинками появляются желтые участки с нормальными щетинками и серые с опаленными щетинками.
Сущность СК:
при митот делении кл, гл.об, в эмбрион период происходят мутации, а затем кроссинговер между двумя сестр хроматидами гомолог хромосом, что приводит к образованию клонов мутантных клеток.
СК проявится только в том случае, если по мутантному аллелю особь гомозиготна.
СК приводит к образованию клонов рекомбинантных клеток, в результате чего, во втором мутационном поколении могут появиться семьи, полностью состоящие из мутантных особей.
Цитологическое док-во кроссинговера.
1931 Эксперименты на дрозофиле, Штерн. Прямые цит.Док-ва кроссинговера.
Мак-Клинтак и Крейтон - опыты на кукурузе (неск позже)
С помощью облучения была получена транслокация Х, У и 4 хромосомы у дрозофилы. В результате у женских особей появились морфологически различные Х-хромосомы (с фрагментом Г-обр хромосомы).
Х-хромосома: гены, отв за цвет и форму глаз (красные и коричневые, круглые и полосковидные глаза)
При анализирующем скрещивании было получено 4 типа потомков, 2 большие группы с сочетанием родит призн и 2 маленькие группы с промежут сочет призн (за счет кроссинговера); у кроссоверных особей обнаружены Г-обр хромосомы.
По количеству кроссоверных особей можно судить о расстоянии между генами.
Генетическое картирование.
Исходя из линейного расположения генов в хромосоме и возможности на основании частоты кроссинговера судить о расстоянии между генами, а, соотв, и их последовательности в хромосоме, составляются генетич карты хромосом.
Генетическое картирование - определение положения гена, как минимум, по отношению к 2 другим.
Генетическая карта хромосомы - план расположения генов в хромосоме.
ГК осуществляется в неск этапов:
1. Определение принадлежность гена к группе сцепления (анализ скрещивание): чем больше генов выявлено ударного вида, тем точнее рез-т картирования.
Спонтанные и индуцированные мутации
2. Все гены разбивают на группы сцепления.
3. Определяется частота перекреста между генами (анализ скрещ)
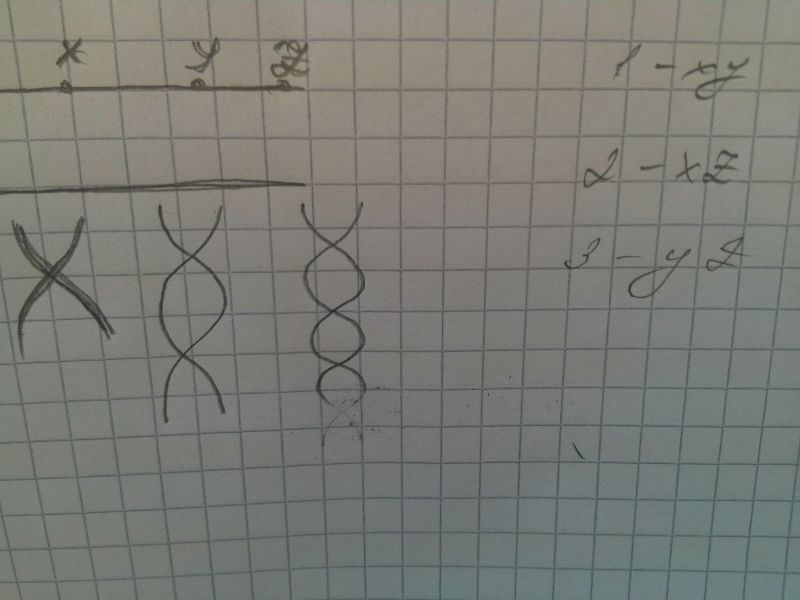
При построении карт указывается расстояние от 0й точки (центромерный, либо теломерный участок)
У бактерий генет карта имеет вид кольца, а расстояние между генами обозначают в ед времени.
При анализе генет карт выявл-ся участки, между кот расст более 50%. Их наличие объясняется суммацией расстояния между очень близкими генами, повторов и молчащей ДНК.
Карта на основании кроссоверных особей - теоретическая/генетическая.
После построения генет карт на основе кроссинговера встает вопрос о соответствии ГК цитологическим, т.е. истинному расположению генов в хромосоме.
Это соответствие впервые было проверено в 1930х гг на гигантских политенных хромосомах.
Каждая хромосома по длине имеет специфический рисунок диска, что позволяет различать участки хромосом. Используя метод хромосомных перестроек, определяют локализацию генов, а затем сравнивают кроссовер ГК с цитолог.
Между ними есть соответствие, но не абсолютное.
Причины несоотв:
1. В хромосоме им-ся области, в которых затруднен или совсем отс кроссинговер (гетерохромат участки)
2. Физическое расстояние увеличивается, если гены разделены молчащей ДНК
Это не снижает ценности ГК. С их помощью можно выявить локализацию дефектных генов и предсказать вероятность появления особей с новым сочетанием признаков.
Хромосомная теория наследственности.
Основные положения.
В основу легли работы Бовери, Сеттона, Моргана и др.
Получены док-ва роли хромосом в передаче наследственной информации.
1. Хромосомы - материальные носители насл инф
2. Элементарной единицей НИ явл-ся ген, локализованный в хромосоме
3. Гены в хромосомах расп-ся в линейном порядке, образуя группы сцепления.
4. Гены, расп-е в одной хромосоме, наследуются совместно
5. Сцепление генов может нарушаться за счет кроссинговера в процессе мейоза
6. В мейозе гомолог хромосомы, а, следовательно, аллельные гены попадают в разные гаметы, которые всегда гаплоидны
7. Негомолог хромосомы расходятся произвольно, независимо друг от друга и образуют различные комбинации в гаметах, число которых 2n, n -кол-во пар гомологичных хромосом.
Наследование пола.
Биологическое значение пола и эволюция раздельнополости.
Формирование пола.
Хромосомный механизм определения пола.
Балансовая теория и половой хроматин.
Наследование признаков сцепленных и ограниченных полом.
Регуляция пола.
Особи разного пола характеризуются различной жизнеспособностью в разные моменты онтогенеза --- соотношение полов 1:1 (примерно)
Пол - совокупность морфологических, физиологических, биохимических, поведенческих и др признаков организма, обусловливающих репродукцию.
Существование 2 полов связано с существованием полового размножения и обусловлено разной ролью м и ж орг-в в половом размножение.
Первичные ПП: органы, принимающие непоср участие в проц воспр-я (пол железы)
Вторичные ПП: органы и свойства, непоср-но не задействованы в репродукции, но способствующие
В начале развития гонады нейтральны --- любой организм бипотенциален.
Если в процессе развития дегенирирует корковый слой первичн пол железы - из мозг в-ва - семенник. Из коркового - яичники.
Становление пола - сложный, многоэтапный процесс, стадии которого строго согласованы. Значение раздельнополости и процесса оплодотворения закл-ся в выгоде, которую получает потомство за счет слияния разнокачественных гамет (Дарвиновское учение).
Эволюционное значение полового процесса:
1. Зигота объединяет различные наследственные задатки и в разнообразныхсочетаниях, т.е половой процесс - источник комбинативной изменчивости, кот является материалом для отбора в любом направлении, содания новых пород и сортов, а также видообразования в дикой природе
2. Сочетание разнокач гамет в зиготе обусловливает высокий жизненный импульс для клеток зародыша, что отражается в высоком уровне обмена и жизненных процессов у нового организма (гетерозис).
Пути детерминации пола:
В ходе эволюции сформ неск механизмов, осн 3:
1. Прогамный (презиготическое определение пола) - пол будущего орг-ма определяется в ходе гаметогенеза у родит особи и различиями размеров гамет. Характерен для коловраток, тлей, некот мор червей и некот растений.
2. Сингамный (зиготическое) - пол определяется в момент слияния гамет. Млекопитающие, птицы, рыбы, двукрылые насекомые, двудомные растения.
3. Эпигамный (постзиготическое) - пол определяется в процессе гаметогенеза и в значит степени зависит от внеш среды. Наступает после оплодотворения. Морской червь бонеллия - если личинка оседает на дно - женская; если свободно плавает, а затем прикрепл-ся к хоботку самки - самец.
4. Эусингамный (гаплоидный). Пчелы, муравьи и др перепончатокрылые
Генотипически пол особи обусловлен различиями в хромосомном аппарате самок и самцов, т.е будет равной вероятность закладки особей м и ж пола.
Пол относят к альтернативным признаком (менделирующий), а соотношение полов 1:1 соответствует расщеплению альтернативного признака в анализирующем скрещивании.
Механизмы определения пола.
Гипотеза зависимости пола от сочетания половых хромосом
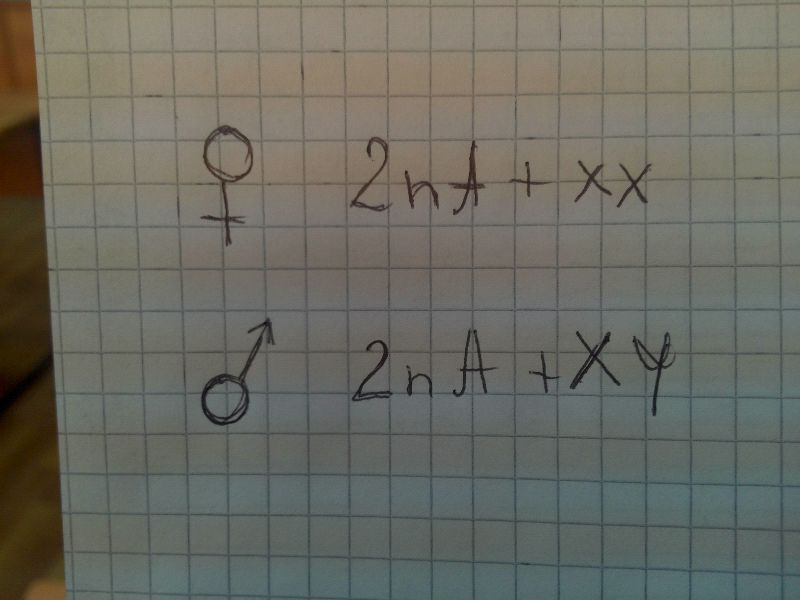
Типы определения пола:
1. Водяной клоп рода Ligaeus, млекопитающие
В сомат клетках самки 2 аутосомы и хх,самца - 2 ауто и ху. Женский организм дает один тип, мужской - 2 типа
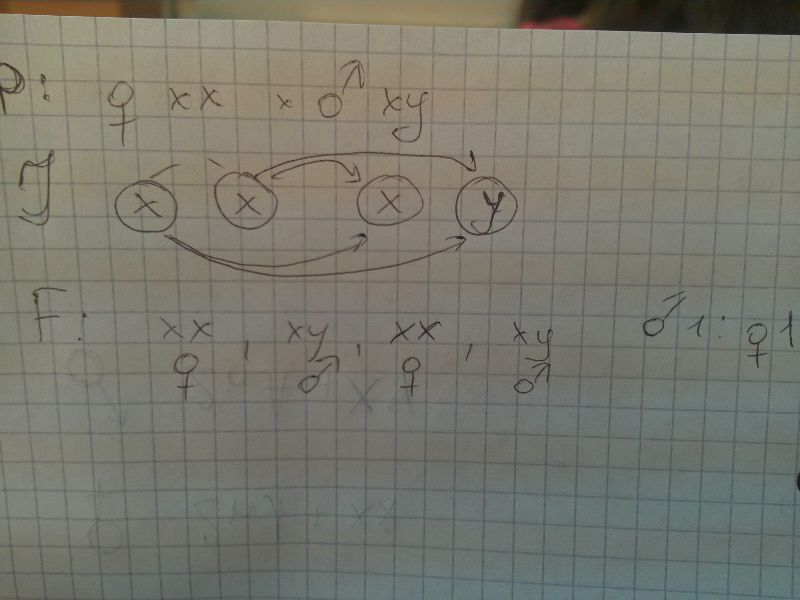
Пол, у которого 2 одинаковые половые хромосомы - гомогаметный, 2 разные - гетерогаметный
2. Водяной клоп Протенор. В сомат клетках самки на 1 хромосому больше, чем у самцов. Ж - гомогаметный, М - гетерогаметный (насекомые)
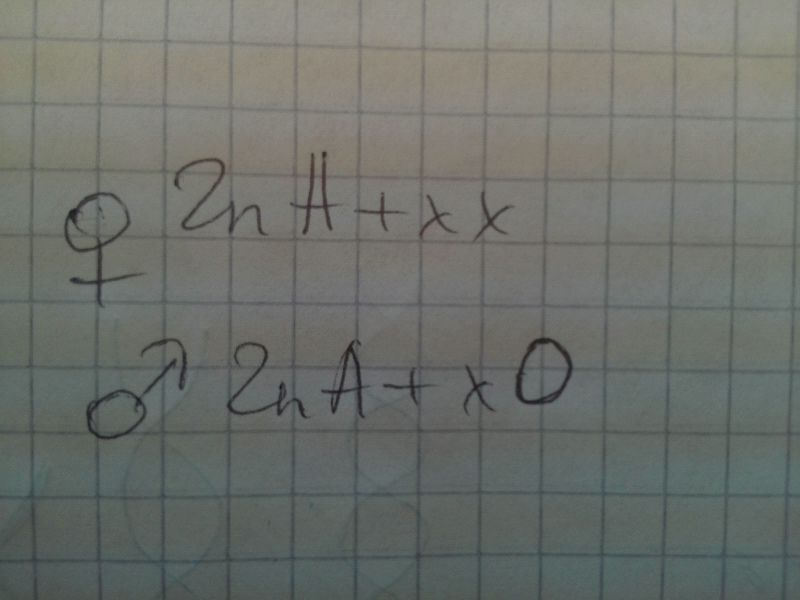
3. Пяденица, вид Абраксас.Ж - гетеро, М - гомо (птицы)

Половые хромосомы Х и У отличаются по размеру и набору генов.
У рептилий пол связан с условиями инкубации яйца. Низкая - самцы, высокая - самки, сочетание - самки, промежут - самцы. Темп интервалы очень малы, пределы кобелания 1-5град.
Пчелы
М - из неоплодотворенных яиц, причем соматич кл способны восстанавливать диплоидный набор, сперматогенез без редукции хромосом.
Ж - диплоидные, 2 типа:
1. Крупные - фертильные (матки)
2. Небольшие (рабочие)
Дифф ж связана с питанием
После проверки хромосом мех-в опред пола, что у некот орг при наличии у-хромосомы, пол мужской --- балансовая теория.
На формирование фенотип проявл пола может более существенно значение оказывать х-хромосома, а у-хромосома на формирование пола не влияет, а оказывает влияние на фертильность пола (дрозофилы): х0 - стерильные м, 2х2у - норм плодовит ж.
После исследований выявилось, что пол в большей степени определяется соотношением половых х-хромосом и аутосом.
в 1925г Бриджис - балансовая теория пола. При нарушении баланса между аутосомами и половыми х-хромосомами получаются формы промежуточного пола - интерсексы. По признакам пола мб сдвинуты в любую сторону.
Закономерность х/А. Если =1 - самки, =0,5 - самцы, >1 &<0,5 - черты интерсексуальности, сверхсамки и сверхсамцы.
Чем больше число аутосом, приходящееся на половые хромосомы, тем сильнее выражен мужской пол. Относительное увеличение половых х-хромосом ведет к образованию сверхсамок (2А+ххх).
Суть БТ:
изначально организм бипотенциален и развитие половых признаков зависит от баланса генов, контролирующих их развитие. Правильность БТ была доказана Бриджисом и Гольдшмиттом при изучении нерасхождения хромосом у дрозофилы и непарного шелкопряда. В рез-те нерасх х-м обр-ся разные гаметы и появляются промежут формы. Т.о форм-ся непрерыв ряд перехода особей от одного пола в другой.
Сверхсамки и сверхсамцы проявляют сильно выраженные признаки того или другого пола и они будут отличаться дозой х-хромосом. Свидетельство: генантроморфы - особи, у кот одна половина тела мужская, а др женская; появляются в результате отставания у-хромосомы в первом делении дробления, потом лизируется. в итоге есть хх - женская половина, х0 - мужская. У дрозофилы - один глаз красный - жен, один белый - муж
17.04.2013 №9