

4. Сучасні методи зоологічних дослідженнь.
На теперішній час в зоологічній науці використовують всі відомі методи біологічних дослідженнь: 1). Найбільш відомий описовий метод чи метод спостереження за об’єктами в живій природі, де вивчають умови існування, зв’язок їх з іншими представниками, вплив на них фізичних та біологічних факторів. Вивчається вплив біотичних та абіотичних факторів на умови розповсюдження тварин. Важливим метод спостереження є при визначенні масової появи корисних та шкідливих тварин, носіїв небезпечних хвороб людини та тварини. Дані метода є найголовнішими при визначенні виду тварини. Інші методи доповнюють та конкретизують попередні дані метода спостереження за об’єктами в природі.
2. Методи експериментальних досліджень: методи анатомічних дослідженнь – використовується при вивченні внутрішніх органів, їх топографію в організмі; методи вивчення макро та мікроструктури за допомогою збільшувальних приладів світлової та електронної мікроскопії; метод гістологічних дослідженнь – використовується при вивченні тканин тварини. Використовуються методи фарбування та ідентифікації тканин. Вивчається внутрішній склад тканин; метод цитологічних дослідженнь – вивчається будова клітин окремих тканин органів;
метод каріологічного аналізу – вивчення будови хромосом, їх особливості; метод біохімічного аналізу – вивчення хімічного складу протоплазми клітин, продуктів гомеостазу; метод молекулярно-біологічного аналізу – вивчення структури хімічних елементів тварини (білки, жири, вуглеводи, нуклеїнові кислоти); використовуються радіобіологічні методи; методи комп’ютерної томографії.
3. Порівняльний метод – дає можливість знайти закономірності, загальні для різних явищ.
4. Статистичний метод – його ще називають біометрією , необхідний для здобуття вичерпної інформації про типові об’єкти, їх різноманіття, про системи біологічних взаємодій та взаємовідношень, вплив різних факторів на біологічні об’єкти.
5. Історичний метод – біологічні об’єкти та явища розглядаються в історичному аспекті: як виникли, розвивалися чи зникли в зв’язку з конкретними умовами.
Значення зоології в життєдіяльності людини, біосфери та охорони навколишнього середовища.
Важко переоцінити те значення, яке має зоологічна наука. Серед числених тварин є чимало цінних для господаства форм, які поставляють людині їжу і сировину для багатьох галузей промисловості. До об’єктів промислу належать ракоподібні, молюски, деякі голкошкірі. Бджоли дають людині мед та віск. Багато хребетних є кормовою базою для промислових риб, наприклад у Каспійському морі акліматизовано багатощетинкового кільчака нереїса, це значно зміцнило кормову базу осетрових.
Безхребетні мають значення в утворенні грунту (дощові черви).
Серед безхребетних тварин є багато форм, які завдають великої шкоди – це комахи, кліщі, ряд нематод. Останнім часом в результаті зменшення пильності за шкідливими комахами зареєстровані напади сарани на півдні України (1999-2000р.)
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 2. Загальна характеристика підцарства одноклітинних.
Питання:
1. Будова найпростіших як одноклітинних організмів.
2. Багатофункціональність клітин найпростіших.
3. Уявлення про органели.
4. Поширення і середовище існування найпростіших.
Клітина найпростішого — це самостійний організм, якому властиві всі життєві функції: обмін речовин, подразливість, розм- ноження, пересування в просторі.
Усі найпростіші належать до еукаріот, їхні клітини мають оформлене ядро, в якому знаходиться генетичний матеріал. Від навколишнього середовища клітина відмежована клітинною мем- браною, яка викопує бар'єрну та захисну функції. У цитоплазмі розрізняють два шари: ектоплазму (зовнішній) і ендоплазму (вну- трішній). До органел, типових для еукаріотичних клітин, належать мітохондрїі, ендоплазматичний ретикулум, рибосоми, апарат Гольджі, лізосоми, ядро. Поряд із ними в цитоплазмі найпростіших присутні специфічні органели — травні та скоротливі вакуолі, ба- зальне тільце (у джгутикових), вічка.
Скоротлива вакуоля — це органела осморегуляції, що кон- тролює надходження води в клітину. Вода накопичується в ско- ротливій вакуолі, а внаслідок її скорочення виливається назовні крізь пору. Інші функції скоротливої вакуолі: виділення — разом з водою назовні виводяться продукти обміну речовин; дихання — з водою надходить розчинений кисень.Травна вакуоля виконує функцію травлення.У її порожнину виділяються ферменти, пере- травлюють поглинені частинки їжі (органічні залишки, бактерії та інш.).
Як і всім живим організмам, найпростішим притаманна по- дразливість — здатність реагувати на різні зміни навколишнього середовища. Подразливість одноклітинних носить характер такси- су. Розрізняють позитивні таксиси — рух до джерела подразнення, і негативні — уникнення дії подразника.
За типом живлення найпростіші можуть бути фототрофами і гетеротрофами. Деякі одноклітинні, наприклад евглена зелена, в умовах яскравого освітлення поводяться як фототрофи, а в темря- ві, за наявності органічних речовин, переходять на гетеротрофний тип живлення. Серед найпростіших зустрічаються вільноживучі (прикріплені, вільноплаваючі) і паразитичні форми. Вільношта- ваючі здатні до активного руху, який забезпечується мінливими виростами клітини — псевдоподіями, (амеби, радіолярії), джгути- ками (хламідомонада, трипаносома), війками (інфузорії). Багато з найпростіших утворюють колонії (вольвокс). Деякі одноклітин- ні пристосувалися до паразитичного способу життя (дизентерійна амеба, малярійний плазмодій).
Найпростіші поширені повсюдно. Вони освоїли прісні, морські води та ґрунт. Багато найпростіших, що живуть у Світовому океа- ні, мають мінеральний скелет (форамініфери, радіолярії) з кальцій карбонату або силіцій оксиду. Після відмирання організмів ці ске- лети утворюють потужні донні поклади.
На сьогодні описано понад ЗО тис. видів найпростіших. Тип Простіших ділять на класи: Саркодові, Джгутикові,Споровики, Інфузорії.
Підтип Саркодові (Sarcodina)
1.Загальна характеристика прісноводних, морських та парзитчних представників.
2.Класифікація.
3.Значення в природі і житті людини.
Клітина саркодових вкрита плазматичною мембраною, проте багато видів мають раковину. Органелами руху і захоплення їжі у представників класу є непостійні вирости цито- плазми — псевдоподії (несправжні ніжки). Основна маса саркодо- вих — мешканці моря; у прісноводних форм є так звані скоротливі вакуолі, що забезпечують виведення з клітини надмірної кількості води. Живляться саркодові водоростями, бактеріями та іншими найпростішими. Розмножуються як нестатевим, так і статевим способом.

Амеба протей — типовий представник саркодових (рис.1). Мешкає в прісних водоймищах. Зовні тіло амеби вкрите плазмалемою. Ха- рактерна особливість — відсутність постійної форми тіла; клітина утворює вирости — псевдоподії (несправжні ніжки), за допомогою яких амеба пересувається. В ендоплазмі розташовується травна вакуоля і численні поживні включення, скоротлива вакуоля, ядро й інші органели. Захоплення їжі здійснюється шляхом фагоцитозу: несправжні ніжки оточують поживну частку, після чого відбувається її поглинання й утворення травної вакуолі.
Клас Джгутикові. Орган руху — джгутики. Більшість джгутикових має постійну форму, проте деякі види зберігають здатність
утворювати псевдоподії. Серед джгутикових є організми з автотрофним, гетеротрофним і змішаним типами живлення.
Евглена зелена (Euglena viridis) характерний представник джгутикових (рис2).

Значно поширена в прісноводних водоймах. Евглена має один джгутик. У передній частині клітини знаходиться яскраво-червоне вічко — стигма, що виконує функцію визначення джерела світла (евглена виявляє позитивний фототаксис). У цитоплазмі присутні зелені хлоропласти, що містять хлорофіл. Зовнішній шар цитоплазми утворює пелікулу.
Евглена здатна змінювати характер живлення й обміну речовин залежно від умов середовища: на світлі їй властивий автотрофний тип, у темряві — гетеротрофний.
Особливості організації представників типу Апікомплексні (Apicomplexa).
1.Особливості організації представників типу в зв’язку з паразитичним способом життя.
2.Життеві цикли розвитку представників.
Клас Споровики (Sporozoa) включає в себе тільки паразитичні форми. Під впливом паразитизму будова споровиків сильно спрощена у порівнянні з представниками інших класів. Вони не мають органоїдів руху, травних та скоротливих вакуолів.
Життєвий цикл відрізняється складністю, проходить зі зміною хазяїв та чергуванням безстатевого, статевого розмноження та спорогонії. Безстатеве розмноження проходить шляхом шизогонії чи множинного поділу. Кінцевою стадією розвитку є утворення спор та спорозоїтів.
Н азва
“споровики”
вказує на нявність у їх життєвому циклі
стадії спори
азва
“споровики”
вказує на нявність у їх життєвому циклі
стадії спори
де зародок паразитів вміщений в особливу оболонку, подібну цистам других простіших. Цим забезпечується розповсюдження та захист зародків від несприятливих зовнішніх умов. Але серед споровиків є такі, що не утворюють спор – це кров’яні споровики. До них відносяться малярійні плазмодяії, що викликають у людей жахливе захворення – малярію.
Малярія, або переміжна лихоманка, — одна з важких і поширених хвороб людини. Так, в Індії число захворювань малярією доходить до декількох мільйонів чоловік в рік, причому хвороба часто закінчується смертю. Хвороба полягає в сильних нападах лихоманки з високою температурою - до 40—41°С. Напади повторюються через дві доби на треті або через троє на четверті, всі більше посилюються і дуже виснажують організм людини. Цю хворобу відвіку називали малярію «болотяною лихоманкою», оскільки частіше за захворювання малярією спостерігалися в місцях, багатих болотами і стоячими водоймищами. Дійсних причин захворювання не знали і тому помилково вважали, що хвороба викликається шкідливими випаровуваннями боліт.
Проте дослідженнями головним чином російських учених були з'ясовані причини, що викликають малярію, а також способи її розповсюдження. Виявилось, що малярію викликає крихітний простий паразит, що потрапляє в кров людини. Зародки малярійного паразита — плазмодія малярії — проникають всередину червоних кров'яних кліток, харчуються ними і ростуть. Плазмодії, що виросли, є маленькою клітинкою, що поміщається усередині червоної кров'яної клітки. Тут же паразити і розмножуються. Клітка малярійного плазмодія розділяється на декілька маленьких клітинок - зародків, які виходять із зруйнованої кров'яної клітки і проникають в інші, здорові кров'яні клітини (рис.3). Під час виходу зародків із зруйнованої кров'яної клітки і відбуваються напади малярії. Це пояснюється тим, що при цьому в кров потрапляють отруйні речовини, що виділяються паразитами. З кожним нападом збільшується число паразитів в крові людини і руйнується вся більша кількість кров'яних кліток. Це веде до сильного недокрів'я і виснаження організму людини.
Відомо, що існують різні форми малярії. У одних випадках напади повторюються через дві доби, в інших — через три доби. Нарешті, існує особливо виснажлива форма малярії - тропічна. Різні форми малярії залежать від зараження людини різними видами плазмодія.
Яким же чином людина заражається малярією і як паразити малярії потрапляють в кров здорової людини? З'ясувалося, що паразити в кров людини попадають за допомогою особливих видів комарів, яких назвали малярійними. Відрізняються від звичайних тим, що на їх крильцях є темні плями, та коли він сідає на тіло людини сисати кров, його черевце підняте вгору (рис.3). Якщо такий комар нассеться крові у людини хворої малярією, то з кров'ю в організм комара потраплять і малярійні паразити. У телі комара на зовнішній оболонці кишки вони утворюють багато зародків - спорозоїтів. Спорозоїти потрапляють в слинні залози комара. Коли заражений комар смокче кров здорової людини, то разом із слиною комара в ранку потрапляють і зародки малярійного паразита. Щорічно від цієї хвороби страждають мільони людей, особливо в теплих країнах Землі – це Індія, північ та центр Африки, Центральна Америка.
Але чому ж малярія поширена в болотистих районах? Це пояснюється тим, що малярійні комарі живуть поблизу болот і стоячих водоймищ, оскільки вони відкладають свої яєчка у воду, з яєчок виводяться личинки. Вони також живуть у воді, але дихають атмосферним -повітрям. Для дихання вони піднімаються до поверхні води (рис.3,2). Личинки, що виросли, обертаються в лялечку, і лялечки (рис.3.3) також живуть у воді до вильоту комарів. Відкриття малярійного паразита і способу його перенесення від хворого до здорової людини малярійними комарами дало можливість вести успішну боротьбу з малярією.
З давних – давен у народі малярія має назву болотної пропасниці, що пов’язане з особливим лютуванням її в болотистих місцевостях, де успішно, як виявили, розвиваються личинки малярійних комарів.
З історії відомо, що все античне Середземномор’є знаходилося в полоні малярії. В самому Римі деякі імператори залишали місто у малярійний сезон року. На території древньої Греції малярія спостошувала цілі міста. За свідченням авторитетних вчених від малярії загинули Олександр Македонський, римський імператор Траян, пророк Мухамед, різні германські середньовікові королі та багато інших.
Одним із засобів боротьби з малярійним комаром є висушування боліт, що дають притулок його личинкам, або отруєння чи нафтування таких водойм.
Характеристика типів Мікроспорідії (Microspora) і Міксоспорідії (Mixozoa).
1.Загальна характеристика.
2.Особливості будови, життєві цикли багатоклітинні фази розвитку. 3.Значення мікроспоридій як паразитів риб. Представник – міксоболюс (Myxobolus cerebralis).
Число відомих видів міксоспорідій близько 1000. Це тканинні або порожнинні паразити, що вражають різноманітні органи господаря: зябра, шкіру, м'язи (мал. 50), жовчний і сечовий міхури, нервову систему, хрящі та ін. Дорослі міксоспорідій, що живуть в порожнинах органів, являють собою амебоподібні завжди багатоядерні плазмодії, розміри яких варіюють від декількох десятків мікрометрів до двох сантиметрів.
Міксоспорідіі локалізуються в товщі тканин і утворюють цисти, пухлини, що досягають 1-2 см (іноді і більше) у діаметрі (рис. 50). Вміст цисти заповнено цитоплазмою паразита з великою кількістю ядер. Останні за структурними і функціональними особливостями поділяються на дві категорії: вегетативні і генеративні. Вегетативні ядра визначають всі життєво необхідні функції плазмодія: обмінні процеси, синтез білків, ріст і ін Генеративні ядра дають початок особливим, службовцям для розмноження стадіям - спорах. Цей процес починається з того, що навколо генеративних ядер відокремлюються невеликі ділянки цитоплазми. Формуються так звані генеративні клітини, звичайно численні і здатні до самостійного пересування всередині тіла плазмодія. Ядро генеративної клітини через деякий час починає ділитися і дає багатоядерні освіта - панспоробласт. Усередині останнього формуються дві суперечки, в побудові кожної з яких бере участь по 6 і більше ядер.
Спори міксоспорідій надзвичайно різноманітні . Зовні вони одягнені міцною оболонкою, яка складається у більшості міксоспорідій з двох з'єднаних щільним швом стулок (ряд Bivalvulea) або (що трапляється значно рідше) з більшого числа стулок, від 3 до 6 (ряд Multivalvulea). Стулки утворюються за рахунок відокремлення невеликих ділянок цитоплазми з одним ядром кожен.
У багатьох міксоспорідій стулки оснащені різними досить довгими виростами . Біологічна роль їх зводиться, ймовірно, до збільшення поверхні, що сприяє пасивному "плаванню" у воді. Усередині утворень формуються жалкі капсули, число яких відповідає числу стулок (від 2 до 6). В утворенні кожної капсули беруть участь ділянки цитоплазми і ядро. Усередині капсули знаходиться згорнута жалка нитка. У порожнині спори розташовується амебоідний зародок з двома ядрами. Таким чином, спору міксоспорідій по суті справи справи багатоклітинне утворення.
Спори з тіла хворої риби потрапляють у воду, заковтуються іншою рибою і в її кишечнику під впливом травного соку жалкі нитки з великою силою викидаються і встромлюють в стінку кишки. Стулки спори розкриваються по шву, амебоїдний зародок виходить із спори, проникає через епітелій кишки в капіляри, і потоком крові доставляється до остаточного місця свого розвитку, де незабаром в результаті ділення ядер перетворюється на багатоядерний плазмодій. В останньому незабаром починається відокремлення генеративних клітин і розвиток спор.
Статевий процес у міксоспорідій відбувається наступним чином. Всі ядра плазмодія (вегетативні та генеративні) диплоїдні. Лише при розвитку спор відбувається мейоз, в результаті чого ядра, формують спори (у тому числі і ядра амебоідного зародка, жалких капсул і стулок), виявляються гаплоїдними. При виході амебоідного зародка з спори (або дещо раніше) ядра зливаються, утворюючи знову диплоидное ядро. До цього й зводиться статевий процес. Подібна форма статевого процесу називається автогамією.
Таким чином, і за характером статевого процесу, і по співвідношенню гаплоїдної і диплоїдної фаз ядра в життєвому циклі міксоспорідії різко відмінні від споровиків, які представляють собою гаплонтів з зі готичною редукцією, всі стадії життєвого циклу, крім зиготи, гаплоїдні. У кнідоспорідій справа йде якраз навпаки - ядра протягом усього циклу диплоїдні і лише ядра амебоідів (які можна порівняти з ядрами гамет) гаплоїдні. Однак питання про форми статевого процесу у міксоспорідій вимагає ще подальших досліджень. Тип війконосні (Ciliophora)
1.Особливості будови (ускладнення органел, живлення і ексреції, війчастий апарат, трихоцисти, ядерний дуалізм) і життєвий цикл. 2.Життєві форми інфузорій: планктонні – інфузорія-туфелька (Paramacium caudatum), трубач (Stentor sp.), придонні – стилоніхія (Stulonichia mutilus), сидячі – сувійка (Vorticella nebulifera), Хижі форми – дідініум (Didinium sp), ряд сисні інфузорії (Suctoria), паразитичні – іхтіофтіріус (Ichthyophthrius sp), тріходіна (Trich0dina sp), ендобіонтні (ряд Ентодінієподібні – Entodiniomorpha) форми. Роль війчастих в природі і господарстві людини (очищення ставків, забруднених вод, біоіндикація).
Клас Інфузорії. Понад 7 тис. видів інфузорій освоїли морські та прісні води, ґрунт, багато з яких пристосувалися до паразитизму.
Інфузорія туфелька (Paramecium coudatum) — одержала свою назву через форму клітини (рис.4). Характерна особливість — наявність безлічі війок по всій поверхні тіла. Війки знаходяться в безперервному русі, що забезпечує
 швидке
переміщення інфузорії в просторі.
швидке
переміщення інфузорії в просторі.
Ектоплазма інфузорії утворює декілька мембран, що надає найпростішому постійної форми тіла. На одному боці у туфельки є заглибина — рот (перистом), у якому розташовані довгі війки, що забезпечують надходження поживних речовин. Кожна з двох скоротливих вакуолей складається з центрального резервуару та декількох привідних канальців, якими в резервуар стікає надмірна кількість рідини. Під час його скорочення рідина виливається назовні. Травна вакуоля інфузорій рухається з потоком цитоплазми. Непсретравлєні залишки їжі викидаються через порошицю, розташовану в певній ділянці тіла.
В інфузорії є два ядра, різні за формою і кількістю генетичного матеріалу. Велике ядро (макронуклеус) містить більше ДНК, характеризується високим рівнем транскрипції, що пов'язано з його участю в процесах синтезу білка. Мале ядро (мікронуклеус) бере участь у процесах розмноження (рис. 4). Розмноження інфузорії може відбуватися як нестатевим способом (поділом) (рис. 5), так і статевим. Зазвичай - статевий процес чергується з декількома циклами нестатевого розмноження.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 3. Основні риси багатоклітинних (Metazoa).
Питання:
1.Основні риси багатоклітинних: клітинна і тваринна диференціація, поділ клітин на соматичні та генеративні, життєвий цикл з гаметичною редукцією.
2.Походження багатоклітинних (гіпотези гастреї, фагоцители, паренхімели) та сучасні погляди на цю проблему.
Будова і функції багатоклітинних організмів. Як вам відомо, у більшості багатоклітинних організмів клітини диференціюються за особливостями будови та функцій, утворюючи різні типи тканин. Різні тканини входять до складу органів. Орган - це певна структура організму, яка складається з тканин різних типів, але, як правило, переважає один із них (наприклад, у серці - м'язова тканина). Кожен орган займає в організмі певне положення, характеризується певними особливостями будови та виконує конкретні функції.
Органи, що виконують спільні функції, утворюють в організмі тварин системи органів, наприклад, дихальну, кровоносну, опорно-ру-хову, сечостатеву та ін. Органи однієї системи можуть послідовно з'єднуватись один з одним (наприклад, органи травної, дихальної систем) або бути «розкиданими» в організмі (ендокринна система).
Органи різних систем можуть тимчасово об'єднуватись для виконання певної функції, утворюючи функціональну систелгу органів (наприклад, під час бігу скоординовано функціонують опорно-рухова, дихальна, кровоносна, нервова системи тощо).
Основними властивостями багатоклітинного організму, як і одноклітинного, є обмін речовин, перетворення енергії, здатність до саморегуляції та розмноження. Будь-який організм є відкритою системою: він потребує постійного надходження ззовні енергетичного та будівельного матеріалу. Особливе місце в організмі належить регуляторним системалі. нервовій, гуморальній та імунній, які забезпечують його функціонування як єдиного цілого, зокрема зумовлюючи певні реакції на зміни умов зовнішнього та внутрішнього середовища.
Основними відмінностями одноклітинних і багатоклітинних організмів є те, що кожен одноклітинний організм виконує всі життєві функції за допомогою органел чи інших клітинних структур, а кожна з клітин багатоклітинних організмів пристосована до виконання лише однієї чи кількох певних функцій у складі певних тканин, які, в свою чергу, утворюють органи. Тому різні прояви життєдіяльності (живлення, дихання, виділення, транспорт речовин, рух, регуляція обміну речовин, розмноження та індивідуальний розвиток) у багатоклітинних організмів лише частково відбуваються на клітинному рівні, а реалізуються здебільшого в тканинах і органах насамперед завдяки взаємодії цих структур. Усі життєві процеси ба гатоклітинних організмів регулюються різноманітними біологічно активними речовинами, а у більшості тварин - ще й нервовою, ендокринною та імунною системами.
Органи багатоклітинних організмів поділяють на вегетативні та генеративні. Перші забезпечують обмін речовин, рух, ріст тощо, другі спеціалізовані для здійснення процесів розмноження.
Багатоклітинні тварини та рослини по-різному реалізують свої життєві функції, що насамперед залежить від способу живлення -гетеротрофного у тварин і автотрофного - у рослин. Проте деякі процеси життєдіяльності цих організмів можуть здійснюватися подібно.
Рослини як автотрофні організми дістають необхідні для процесів біосинтезу речовини з грунту (розчини мінеральних солей) та повітря (вуглекислий газ), а необхідну енергію - від сонячного проміння. Рослини, на відміну від тварин, ведуть переважно прикріпле-ний спосіб життя, у них відсутні нервова система, органи чуттів, спеціалізовані травна, дихальна, видільна системи тощо. Провідні тканини забезпечують транспорт води, розчинів мінеральних та органічних сполук, біологічно активних регуляторних речовин.
Багатоклітинні тварини як гетеротрофи активно використовують різні джерела живлення, багаті на органічні сполуки. Вони мають органи чуттів, нервову та опорно-рухову системи, структури для захоплення та перероблення їжі (травна система). Це сприяло інтенсифікації обміну речовин і перетворенню енергії, забезпечило активний спосіб життя, а у теплокровних тварин (птахи, ссавці) зникла залежність температури тіла від умов довкілля.
У тварин є особливі системи ефективного транспорту кисню до окремих тканин і клітин (дихальна та кровоносна) та розподілу речовин між різними частинами організму (лімфатична і кровоносна системи, порожнинні рідини), а також спеціальні органи виділення, які, крім виведення з організму продуктів обміну, водночас беруть участь у підтриманні гомеостазу.
Характеристика Типу Губок (Spongia)
Основні відмінності в будові губок.
Типи клітин стінки тіла губок.
Розмноження, розвиток, поширення губок.
Губки — примітивніші багатоклітинні тварини, ведуть прикріплений спосіб життя. Перші представники губок з'явилися в протерозойську еру. На сьогодні відомо понад 3000 видів губок. Губки поширені в прісних і солоних водах усіх кліматичних зон, представлені як поодинокими, так і колоніальними формами.
Незважаючи на таку різноманітність, усі губки мають загальний план будови, що дозволяв об'єднати їх в один тип:
клітини тіла диференційовані та мають тенденцію до утворення тканин;
тіло складаеться з двохшарів клггин — ектодерми и ентодерми, між ними знаходиться драглиста речовина — мезоглея;
3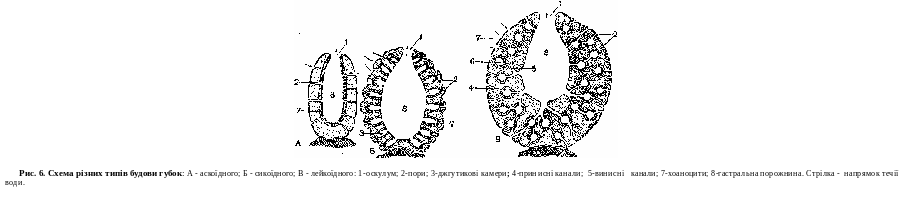 )
майже
завжди є внутрішній скелет (вапняковий,
кремнієвий), який виконує опорну функцію
(рис.6).
)
майже
завжди є внутрішній скелет (вапняковий,
кремнієвий), який виконує опорну функцію
(рис.6).
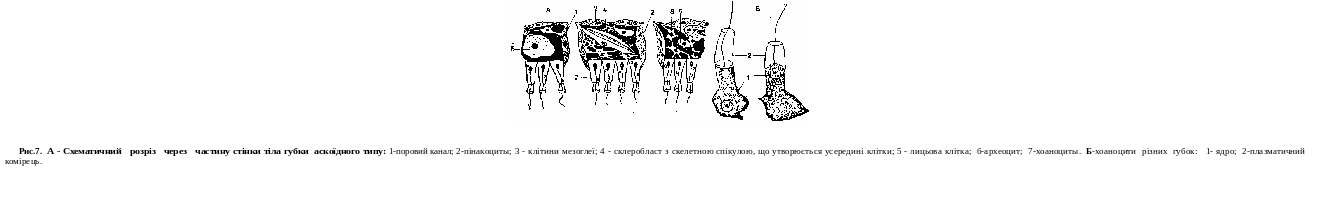
У багатьох губок тіло має вигляд келиха або мішечка, прикріпленого до субстрату (дна, камінця, раковин). У верхній частині знаходиться отвір — гирло, через яке порожнина губки (атріальна порожнина) сполучаеться з навколишнім середовищем. Стінка тіла складаеться з двох шарів — екто- і ентодерми. В ектодермі знаходяться плоекі клітини, які утворюють покривний опітелій.
Ентодерму складаютъ клітини, наділені джгутиком,— хоаноцисти. У мезоглею занурені: опорні клітини; клігини, що формують скелет; амебоцити, що мають псевдоподії, які беруть участь у травленні, здатні перетворюватися на інші види кл!тин; статеві клітини. Стін- ка тіла губки пронизана численними наскрізними порами, в яких знаходяться хоаноцити. Скелет складаеться з безлічі голок (спікул), що мають різноманітну форму и розміри. В утворенні скелета боре участь спонгін — речовина, що скріплює голки між собою (рис.6).
Живлення, дихання і виділення здійснюються за допомогою безперервного потоку води крізь тіло. Завдяки ритмічній роботі джгутиків хоаноцистів, вода нагнітаеться в пори. Потраляє в атріальну порожнину і через гирло виводиться назовні. Залишки відмерлих організмів і найпростіші, які містяться у воді, захоплюються хоаноцитами, передаються амебоцитам і розносяться ними по всьому тілу. Захоплення поживних частинок відбувається шляхом фагоцитозу, травлення у губок внутрішньо-клітинне. Неперетравлені залишки викидаються в порожнину і виводяться назовні. Для дихання використовується розчинений у воді кисень, який поглинається всіма клітинами тіла. Вуглекислий газ також виводитьсяв розчиненому стані.
Розмножуються губки як статевим, так і нестатевим шляхом.
Головною причиною, яка перешкоджає масовому розповсюдженню губок, є відсутність відповідного субстрату. Більшість губок не може жити на мулистому дні, оскільки частинки мулу закупорюють пори, що призводить до загибелі тварини. Великий вплив на поширення мають солоність і рухливість води, температура.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 4. Загальна характеристика підрозділу
Променеві (Radiata)
Тип кишковопорожнинні (Coelenterata).
Питання:
Загальна характеристика типу;
Особливості будови прісноводної гідри. Розмноженя. Регенерація.
Будова гідроідної та сцифоїдної медузи. Розмноження.
Будова коралових поліпів. Значення.
Кишковопорожнинні — примітивні багатоклітинні водні організми, що виникли в протерозойську еру. Відомо до 9 тис. видів, серед яких є вільноживучі, прикріплені та колоніальні форми. Усім представникам типу властиві такі особливості:
тіло складається з двох шарів — екто- й ентодерми, між якими є мезоглея;
в ектодермі знаходяться жалкі клітини, отрута яких паралізує жертву й відлякує ворогів;
тіло має променеву симетрію;
вперше з'являється нервова система;
намічається тенденція до порожнинного травлення, яке відбувається в гастральній (кишковій) порожнині.
Унікальною особливістю кишковопорожнинних є наявність жалких клітин. Кожна жалка клітина має капсулу, заповнену отрутою; у капсулу занурена згорнута жалка нитка. На поверхні клітини знаходиться чутлива волосина, уразі дотику до якої жалка нитка, рясно змочена отрутою, викидається назовні та встромлюється в тіло жертви. Отрута кишковопорожнинних має нервово-паралітичну дію.
У типі Кишковопорожнинні виділяють три класи: Гідроїдні, Сцифоїдні, Коралові поліпи.
Клас Гідроїдні. Клас гідроїдних складається з найбільш примітивних кишковопорожнинних. Серед них є колоніальні і поодинокі форми. Живуть вони і в морській і в прісній воді. До прісноводних форм належить прісноводна гідра. Гідра — це типовий поліп, що має видовжену форму невеликих розмірів (до 1см. завдовжки) (рис.7.А).

На передньому кінці гідри знаходиться ротовий отвір, оточений віночком із 5—12 щупалець, на задньому — підошва, за допомогою якої тварина прикріплюється до субстрату. Стінка тіла складається з екто і ентодерми, а між ними знаходиться драглиста маса — мезоглея. Ектодерма утворена епітеліально-м'язовими клітинами, в яких розрізняють тіло (виконує покривну функцію) і довгий відросток. Відросток має м'язове волокно; сукупність відростків дає можливість щупальцям і тілу гідри стискатися у разі дії подразників. В ектодермі також знаходяться жалкі, нервові, статеві і недиференційовані клітини. Нервові клітини мають довгі відростки, які утворюють у сукупності нервове плетиво. Така нервова система називається дифузною. Ентодерма представлена двома типами клітин: епітеліально-м'язовими і залозистими. Перші, як і відповідні клітини ектодерми, мають відростки і також беруть участь у рухах гідри. Крім того, вони несуть 2—5 тонких джгутиків, спрямованих у кишкову порожнину, які виконують важливу роль у процесах живлення (рис.7.Б).
Гідра, як і інші кишковопорожнинні, — хижак; живиться дрібними водними тваринами, личинками комах, інфузоріями.
Гідри — малорухливі тварини; більшу частину часу проводять у прикріпленому стані. Пересуваються вони, почергово прикріплюючись до поверхні то ротовим кінцем тіла, та підошвою.
П рісноводна
гідра розмножується нестатевим
(вегетативним)
і
статевим шляхом. Нестатеве розмноження
відбувається звичайно брунькуванням
рідко поздовжнім або поперечним поділом.
Бруньки утворюються приблизно на
середині тіла в так званому поясі
брунькування. У цьому поясі час від часу
утворюється горбочок, який поступово
збульшується, на його вільному кінці
виростають щупальця і утворюється рот.
У своїй основі така молода гідра поступово
відшнуровується, відривається від
материнського організму і починає жити
самостійно. Способом брунькування гідра
розмножується при нормальних умовах
протягом літа, а восени розвиваються
статеві клітини (рис.8.В).
рісноводна
гідра розмножується нестатевим
(вегетативним)
і
статевим шляхом. Нестатеве розмноження
відбувається звичайно брунькуванням
рідко поздовжнім або поперечним поділом.
Бруньки утворюються приблизно на
середині тіла в так званому поясі
брунькування. У цьому поясі час від часу
утворюється горбочок, який поступово
збульшується, на його вільному кінці
виростають щупальця і утворюється рот.
У своїй основі така молода гідра поступово
відшнуровується, відривається від
материнського організму і починає жити
самостійно. Способом брунькування гідра
розмножується при нормальних умовах
протягом літа, а восени розвиваються
статеві клітини (рис.8.В).
Більшість гідр гермафродити. Сперматозоїти і яйцеклітини утворюються у них в ектодермі, причому сперматозоїдні горбки розміщуються блище до ротового полюса, а в горбках – блище до основи формується по одній яйцевій клітині. Зрілі сперматозоїди виходять у воду і проникають в яйцеву клітину (рис.8.А). Так утворюється запліднене яйце. Воно покривається щільною оболонкою. Незабаром гідра гине, а захищені оболонками яйця зимують у водоймищі. Весною з них розвиваються маленькі гідрочки. Таким чином відбувається статеве розмноження гідр. При цьому молодий організм розвивається з однієї заплідненої яєчної клітки. Це служить одним з доказів того, що багатоклітинні тварини походять від одноклітинних.
Більшість морських гідроїдних поліпів утворюють колонії, що складаються з великої кількості особин. Колонія найчастіше має вигляд розгалуженої рослинки (рис.9).
Морські
гідрозої розмножуються як статевим,
так і нестатевим шляхом. Гілочки колонії
утворюються лише шляхом брунькування.
Таким чином утворюється комплекс
поліпів, що сидять неначе на стовбурі
та його гілочках.Статеві клітини
формуються в ектодермі окремих статевих
особинах, що виникають на колоніях також
способом брунькування і називаються
медузами. В ектодермі медуз, що мають
статеві залози розвиваються, розвиваються
сперматозоїти та яйцеклітини. Статеві
клітини виходять у воду де відбувається
запліднення. З заплідненого яйця виходить
личинка – планула, яка виконує роль
розселення організмів. Згодом вона
опускається на дно і прикріплюється до
нього своїм розширеним кінцем. Поступово
відбувається розвиток поліпа, який
розмножуючись безстатевим шляхом формує
нову колонію (рис.9).
Клас Сцифоїдні (Scyphozoa)
Питання:
1.Особливості організації та життєвого циклу на прикладі морської тарілки (Aurelia aurita).
2.Інші представники класу – коренерот (Phizostoma pulmo), люцернарія дзвоникова (Lucemaria campanulata).
3.Отруйні. Засоби лікування при враженні медузою.
Сцифомедузи – виключно морські тварини різного розміру – від декількох сантиметрів - до 2м у діаметрі купола та довжиною щупалець до 30м (рис.10). Тіло сцифоїдної медузи (купол) має форму парасольки або дзвона. У центрі нижнього боку купола знаходиться ротовий отвір, по краях — щупальця, рясно вкриті жалкими клітинами. Деякі щупальця медуз видозмінюються, сильно зменшуються в розмірах і несуть органи чуттів — вічка (світлочутливі органи) і статоцисти (органи рівноваги).
Нервова система сцифоїдних дифузного типу. У ділянці щупалець спостерігається концентрація нервових клітин.
Рот веде до великої гастральної порожнини, яка має чотири бічні вирости (радіальні канали), які часто відкриваються в замкнений кільцевий канал, що йде по периферії парасольки.
Б ільшість
сцифомедуз живляться різноманітними
тваринами планктону: черв’яками,
рачками, дрібною рибою, - коренероті
медузи живляться мікроскопчними
їстивними часточками, що втягують разом
з водою.
ільшість
сцифомедуз живляться різноманітними
тваринами планктону: черв’яками,
рачками, дрібною рибою, - коренероті
медузи живляться мікроскопчними
їстивними часточками, що втягують разом
з водою.
Розмноження і розвиток. Сцифомедузи здебільшого різностатеві. Статеві залози формуються під радіальними каналами другого порядку в ентодермі (на відміну від гідромедуз, в яких статеві продукти утворюються в ектодермі). Дозрілі статеві клітини виводяться назовні через рот, прориваючи стінки радіальних каналів. Запліднення відбувається у воді. В результаті повного і рівномірного дроблення утворюється типова личинка - миготлива планула. Деякий час така личинка вільно плаває, потім опускається на дно, до якого прикріплюється переднім кінцем і перетворюється в сцифістому, що має поліпоїдну форму. Поліпи сцифоїдних ведуть прикріплений спосіб життя і не утворюють колоній. Сцифістома має здатність до бокового і кільцевого брунькування. Внаслідок бокового брунькування утворюються нові сцифістоми, але головним е кільцеве брунькування, або стробіляція, в результаті чого утворюються медузи. Молоді медузи, що відірвалися від стробіли, називаються ефірами. Від дорослої вони відрізняються деяким спрощенням будови – статевих залоз немає, радіальних каналів буває лише чотири. Вони рухливі, активно живляться, ростуть і незабаром перетворюються на дорослих медуз.
Отже, у сцифомедуз добре виражене чергування поколінь. Медузоїдна стадія у них є основною стадією, а поліпоїдна (у вигляді сцифістоми) тільки тимчасова і короткотермінова.
Сцифомедузи рухаються за допомогою досить частих скорочень дзвона (до 140 за хвилину).
Медузи живуть у різних температурних умовах верхніх шарів води, але є глибоководні форми. Деякі сцифомедузи споживаються і служать об’єктами промислу.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 5. Клас коралові поліпи (Anthozoa).
Питання:
1.Загальна характеристика, як морських придонних тварин.
2.Поодинокі корали з ряду актиній (актинія кінська – Actinia eguina). 3.Колоніальні форми – мадрепорові (Scleractinia) корали та ін.. Коралові споруди (берегові, бар’єрні рифи та атоли), як особливості високопродуктивні екосистеми. Ч.Дарвін про утворення атолів.
3.Викопні кишковопорожнинні. Філогенія кишковопорожнинних.
До класу коралових поліпів належать виключно морські твлрини, що мають лише поліпоїдну форму. Переважна більшість коралових поліпів веде сидячий спосіб життя та живе колоніями, які можуть досягати великих розмірів. Симетрія тіла коралових поліпів або восьми-променева, або кількість променів більша і кратна шести. Проте мають місце і випадки більш спеціалізованої, так званої двопроменевої симетрії, що наближається до білатеральної симетрії вищих тварин.
Тіло коралових поліпів має форму циліндричного мішка, одним кінцем якого (підошвою) тварина прикріплюється до субстрату, а на про тилежному кінці утворюється ротовий отвір, який має форму щілини з широким жолобком (с и ф о н о г л і ф). Подібно до гідроїдних поліпів ротовий отвір їх оточений віночком порожнистих щупальців, розміщених біля переходу ротової щілини в стінку тіла. Щупальця можуть мати просту або перисту будову. Порожнина щупальців, сполучається з гастральною порожниною.
Число щупальців відповідно до симетричної будови в одних форм дорівнює восьми, а в інших кратне шести.
Від гідроїдних поліпів коралові поліпи відрізняються значно вищою організацією. Стінка їх тіла має значно складнішу будову, ніж у гідрозоїв. Тут ми зустрічаємось з наявністю самостійних поздовжніх і кільцевих м'язових клітин, що лежать під ектодермою та ентодермою. Нервове сплетення також більш розвинуте. Воно більш заглиблене в стінці тіла і містить у собі чутливі й ган-гліозні клітини. Більш розвинута і мезоглея, що має драглистий або волокнистий вигляд з зануреними в неї клітинами, які походять від ектодерми і ентодерми.
Значно складніша тут і гастральна система. Ротовий отвір веде у трубку (глотку), що звисає в гастральну порожнину. Глотка встелена ектодермою, що з країв ротового отвору загортається всередину. Гастральна порожнина поділена перегородками, або септими, які відходять від внутрішньої поверхні стінки тіла і поділяють гастральну порожнину на камери. У верхньому кінці септи зростаються своїми краями з, глоткою, а нижче виступають у гастральну порожнину вільними краями. Вільні краї септ потовщені і мають вигляд звивистого утвору, що називається мезеннтеріальним шнуром. В епітелії мезентеріальних шнурів містяться травні залози, що беруть участь у травленні. На нижньому кінці шнура є велика кількість жалких клітин, причому цей кінець у деяких форм буває досить довгим і може викидатись через рот або спеціальні отвори. Шнур служить для захисту і нападу. Такі нитковидні утвори називаються аконціями. Коралові поліпи, за винятком деяких поодиноких форм (наприклад, актиній), мають скелет, який найчастіше складається з вапна, рідше — з рогової речовини, а в деяких форм — з того й іншого. Скелет може бути або зовнішній, або внутрішній (рис.11). Розмноження у коралових поліпів буває статеве і нестатеве. У колоніальних форм нестатеве розмноження відбувається способом брунькування, а у поодиноких форм спостерігається поздовжній або поперечний поділ. Статеві клітини визрівають в ентодермі. Запліднення і розвиток яйця відбувається в гастральній порожнині аж до утворення планули, яка через рот виходить назовні, деякий час вільно плаває, а потім осідає на дно, прикріплюється переднім кінцем до нього і перетворюється на поліп.
К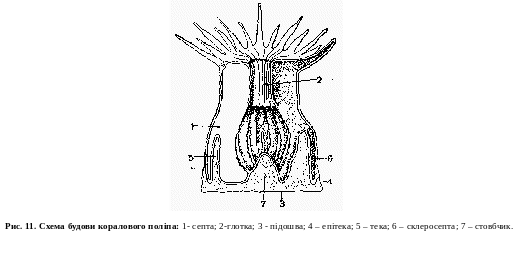 оралові
поліпи роздільностатеві. Сперматозоїди
виходять назовні через рот, прориваючи
стінку тіла, і через рот проникають в
особину іншої статі. Коралові поліпи
поділяються на два підкласи: восьми-променеві
і шести-променеві
корали.
оралові
поліпи роздільностатеві. Сперматозоїди
виходять назовні через рот, прориваючи
стінку тіла, і через рот проникають в
особину іншої статі. Коралові поліпи
поділяються на два підкласи: восьми-променеві
і шести-променеві
корали.
З колоніальних форм слід виділити каменясті чи мадрепорові корали. Їх клонії складаються з незчислених кількостей поліпів, що мають вапняковий скелет. При розростанні таких скелетів утворюються коралові рифи та острова. Біля східного уберіжжя Австралії є величезний бар’єрний риф довжиною 1400км. Коралові острови, чи атоли, мають вигляд кільця; всередині яких знаходиться лагуна. Коралові скелети мають різні кольорові відтінки блідно-рожевого чи червоного кольору. Ці види коралів використовуються людиною для виготовлення прикрас.
Тип Реброплави або гребінники (Ctenophora)
1.Загальна характеристика,
2.Особливості будови, життєдіяльності та розвитку представників класу.
(Ctenophora) — тип морських безхребетних тварин. Тіло (довж. від 2 мм до 1,5 м) найчастіше мішковидне, радіально-симетричне (двопроменеве), драглисте, прозоре. Чисто зовнішню схожість реброплавів з медузами довгий час служила причиною того, що зоологи вважали їх медузами або тваринами, дуже близько спорідненими медузам. На відміну від кишковопорожнинних, до яких Р. відносили на правах підтипу, Р. не мають жалких клітин (див. Кнідобласти). У більшості Р.— 2 щупальця з клейкими клітинами, за допомогою яких Р. захоплюють їжу (дрібні організми планктону, ікра, іноді мальки риб та ін.). P.— вільноплаваючі, рідше повзаючі або сидячі (прикріплені) тварини. Пересуваються за допомогою 8 рядів меридіонально розташованих гребних пластинок (з'єднаних між собою війок), що сидять на потовщеннях тіла — т. з. ребрах (звідси й назва типу). Р.— гермафродити (див. Гермафродитизм), розмноження статеве. Життєвий цикл без метаморфозу. Тип включає єдиний клас з 6 рядами, що об'єднують бл. 90 видів, поширених у всіх морях. Зустрічаються гол. чин. в пн. (4 види) і далекосхідних морях. У Чорному м. 1 вид — Pleurobrachia rhododactyla.
Перші згадки про реброплава можна знайти у давньогрецького вченого Аристотеля, який жив у V-IV ст. до нашої ери. Однак у ті часи зоологічні пізнання були ще дуже примітивні, і Аристотель змішував реброплавів з голотурія, що відносяться до зовсім іншого типу тварин - голкошкірих. Тільки в 1671 р. Мартенс (Martens) у своїй книзі про тварин, які населяють море поблизу Шпіцбергена, привів перший опис і малюнок реброплава. Лінней, творець першої наукової системи органічного світу, приділив реброплава дуже невелику увагу, згадавши в своїй «Системі природи» один вид реброплавів. У 1829 р. вийшла в світ перша у світі велика робота, присвячена медузам. Її автор, німецький зоолог Ешшольц (Eschscholtz), описав в ній і кілька видів відомих йому реброплавів. Він вважав їх особливим класом медуз, який назвав реброплава (Ctenophora). Ця назва збереглася за ними і в даний час. Пізніше з'явилося кілька великих досліджень, присвячених вивченню реброплавів. З них необхідно відзначити книгу Карла Xуна (Chun, 1880), в якій детально описана анатомія середземноморських реброплавів, а також роботи А. О. Ковалевського, І. І. Мечникова та О. Коротнева.
Докладні відомості про реброплава наведені в німецькому керівництві по зоології. Автор розділу про реброплава - проф. Крумбах (Krumbach, 1925). Тим не менш і зараз ця група тварин залишається ще слабо вивченою; особливо мало було відомо про спосіб життя і поведінку реброплавів. Тільки в самий останній час завдяки працям радянського зоолога проф. М. М. Камшилова багато важливих і дуже цікаві особливості біології реброплавів отримали пояснення. Реброплави виключно морські, переважно свободноплавающіе організми. Зазвичай тіло реброплава округлої або мішкоподібні форми. На одному кінці міститься рот. По поверхні тіла реброплава в меридіональному напрямку проходить вісім рядів гребних пластинок. Кожна пластинка розщеплена по зовнішньому краю і має вигляд гребеня, за що гребневики і отримали свою назву. Кожна веслова пластинка складається з ряду злилися між собою великих війок епітелію. Платівки б'ють по воді і, діючи на зразок веселий, повільно пересувають тварина ротовим кінцем уперед.
Більшість видів реброплавів має щупальця. Щупалець завжди два, і розташовуються вони з боків тіла, причому можуть втягуватися в спеціальні щупальцевих кишені.
Щупальця здатні далеко витягуватися і зазвичай перевищують довжину тваринного у багато разів. Від однієї зі сторін щупальця відходять ниткоподібні відгалуження. Вся поверхня щупальця і u200bu200bйого відгалужень покрита численними клейкими клітинами. Окрема клітина має вигляд півкулі, обсаджений липкими сосочками. Під мікроскопом така клітина зовні нагадує за будовою ягоду малини. Від заснування півкулі всередину щупальця відходить спірально закручена нитка, прикріплена своїм іншим кінцем до поздовжнього пучка м'язів, що проходять і вздовж щупальця, і вздовж його гілок. Дрібні морські організми при зіткненні з щупальцями виявляються приклеєними до них липкими півкулями. Якщо видобуток намагається вирватися, то липкі клітини відокремлюються від поверхні щупальця, але залишаються з'єднаними з ним за допомогою еластичної спіральної нитки. Жертва міцно утримується численними клейкими клітинами, нитки яких служать своєрідними пружинками, пом'якшуючими ривки спійманого жівотного.Подобним чином за допомогою еластичного вудилища рибалка утримує на тонкій волосіні велику рибу. Заплуталася в щупальцях видобуток гребневик захоплює краями рота і поїдає. Рот веде в трубковідную глотку, яка в свою чергу відкривається у шлунок. Від шлунка відходять канали гастроваскулярной системи. Один канал продовжується в бік, протилежний роті, і тут розділяється на чотири коротких канальця, два з яких закінчуються сліпо, а два інших відкриваються назовні порами. У сторони від шлунка відходять два канали, які незабаром двічі розгалужуються, завдяки чому до краях тіла проходить вже вісім каналів. Кожен з них впадає в один з восьми меридіональних каналів, що проходять підрядами гребних пластинок. Меридіональні канали тягнуться від ротового кінця тіла до протилежного і сліпо закінчуються на обох кінцях. За ним поживні речовини надходять до місця їх найбільш інтенсивного витрачання - до гребним пластинках.
Нервова система реброплавів дуже примітивна, вона складається з підшкірного нервового сплетення, що має вигляд мережі, і невеликого, але досить густого скупчення гангліозних клітин на аборальной (протилежної ротового отвору) стороні тіла. Тут же знаходиться складно влаштований, так званий аборальний, орган, який, мабуть, грає роль органу рівноваги і регулює інтенсивність роботи окремих рядів гребних пластинок. Основу аборального органу складають чотири хрестоподібно розташовані еластичні дужки, що утворилися з довгих, що злилися між собою вій. На дужках підвішений статоліт-кулька, що складається з вапняних зерен. Під статоліти стінка тіла утворює впячивание, причому ектодерма тут стовщена як подушечки. Зовні статоліт прикритий конічним шатром з злиплих довгих вій. Підстави дужок, що утримують статоліт, за допомогою особливих борозенок, покритих миготливим епітелієм, повідомляються з рядами гребних пластинок. Висить на пружних дужках статоліт безперервно коливається, його руху через дужки і миготливі борозенки передаються рядах гребних пластинок, викликаючи в них хвилю биття, що йде в напрямку від аборального органу до ротового кінця тіла. Якщо видалити аборальний орган, руху гребних пластинок порушуються.
Нещодавно (Камшілов, 1964) було встановлено, що гребневики здатні відрізняти світло від темряви, хоча у них ще не були виявлені спеціальні світлочутливі органи.
На світлі тіло реброплава бероє (Вегое) має жовтувато-рожевий колір. Забарвлення обумовлена u200bu200bнаявністю в покривах тваринного маленьких клітин, наповнених барвником. Ці клітини здатні розтягуватися і стискатися. Якщо помістити бероє у темряву, то вже через 12 - 15 хвилин фарбувальні клітини (хроматофори) починають скорочуватися. Через годину тварина стає молочно-білим. На світлі колір швидко відновлюється і вже через 6-7 хвилин гребневик набуває первісну жовтувато-рожеве забарвлення.
У реброплавів дуже сильно розвинена мезоглея, що надає їх тіла значну прозорість. Зазвичай гребневики безбарвні, тільки деякі з них, як згаданий вище бероє, мають жовтувато-рожеве забарвлення, лише ряди рухомих гребних пластинок переливають всіма кольорами веселки.
Гребневпкі розмножуються виключно статевим шляхом, причому одна тварина утворює і яйця і спермін. Статеві залози мають вигляд довгих колбасовідний тіл, розташованих уздовж меридіональних каналів. З одного боку каналу лежить семенник, з іншого - яєчник. Таким чином, ктенофора має 8 жіночих і 8 чоловічих статевих залоз, не пов'язаних один з одним. Коли статеві клітини дозрівають, стінка, що відокремлює статеву залозу від меридіонального капала, проривається, після чого статеві продукти через просвіт каналу і ротовий отвір потрапляють у морську воду, де і відбувається їх розвиток.
У процесі дроблення яйця утворюються клітини неоднакової величини. На одному полюсі виникає велике число швидко діляться дрібних клітин, які згодом утворюють ектодерми, на іншому полюсі клітини більший і діляться більш повільно. З них утворюється ентодерма майбутнього тварини. Ротовий отвір і гастральная порожнину виникають внаслідок впячивания прошарку великих клітин всередину. Впячивается також і частину дрібних клітин, з яких формується згодом ковтка. Дуже важливо відзначити, що частина великих клітин з глибини гастральной порожнини зародка занурюється в простір між ектодерми і ентодерми. Тут з них згодом формується мускулатура. Ці клітини є зачаток третього зародкового шару - мезодерми. В даний час відомо близько 80 видів реброплавів. Найменші з них, наприклад чорноморська плевробрахія (Pleurobrachia pileus), ледь сягає 5 мм у довжину (не рахуючи щупалець, які можуть витягуватися дуже значно). Найбільш великі види, такі, як венерин пояс (Cestus veneris), бувають до 1, 5 м завдовжки.
Більшість реброплавів живе у товщі води. Через ніжності їх тканин вони вважають за краще триматися в спокійній воді, під час хвилювання опускаються в глибину. Окремі види зустрічаються на глибині 3 км. Багато гребневики в нічний час випромінюють яскраве блакитне світло. Існує припущення, що світяться не самі гребневики, а живуть в їх кишечнику бактерії.
Всі гребневики (за одним винятком) - хижаки, причому до недавнього часу вважалося, що вони харчуються одними тільки планктонними рачками і є конкурентами оселедця, сардини та інших планктоноядних риб. Тому всіх без винятку реброплавів відносили до шкідливих тварин, і дуже часто відсутність в окремих ділянках моря планктоноядних риб пояснювали тим, що гребневики виїли всю їхню їжу. Це здавалося тим більш правдоподібним, що гребневики дуже швидко розмножуються і зустрічаються в масових кількостях. Такі великі гребневики, як болінопсіс (Bolinopsis infundibulim) і бероє (Beroe cucumis), 12 - 15 см завдовжки, за один раз виметивают від 1500 до 3 тис. яєць. При цьому молодь швидко зростає, досягаючи за місяць довжини 8-10 см, верб свою чергу приступає до розмноження.
Вивчення життя реброплавів показало, що кожен з них має свої особливі пристосування для лову видобутку і що різні види реброплавів харчуються різними тваринами.
Реброплави, забезпечені щупальцями, ловлять свою здобич - дрібних планктонних рачків - за допомогою клейких клітин. Болінопсіс, у якого немає щупалець, має по краях рота дві великі лопаті. Розмахуючи ними у воді, болінопсіс заганяє в рот дрібні планктонні організми. У шлунку цього виду були виявлені личинки кільчастих хробаків, різні планктонні рачки, личинки молюсків та інші дрібні планктонні організми. Ці гребневики, безсумнівно, є конкурентами планктоноядних риб. Довгий час за аналогією бероє також вважався шкідливим тваринам, виїдають планктон і нападаючим навіть на мальків риб.
Проте така думка не було підкріплено фактичними даними. Вивчаючи біологію реброплавів, проф. М. М. Камшілов звернув увагу на одну дуже важливу обставину в будові бероє. Цей гребневик малорухомий і позбавлений ловчих щупалець і лопатей. Як же він ловить швидких і хитроверхих рачків, а тим більше рибок? Вивчення вмісту шлунка бероє не дало ніяких результатів - залишків рачків виявлено не було. Тоді стали пропонувати бероє, вміщеним в акваріум, саму різну їжу. Але знову невдача - гребневики відмовлялися від запропонованих їм рачків та інших планктонних організмів. Більше того, виявилося, що, якщо штучно ввести рачка в шлунок бероє, він незабаром викидає його через рот, причому рачок продовжує спокійно плавати - травні соки бероє на нього не діють.
В одному з дослідів в посудину, де містився бероє, був поміщений гребневик болінопсіс. Тоді поводження бероє різко змінилося, він не поспішаючи підплив до болінопсісу, широко розкрив рот і проковтнув реброплава, величина якого була майже такою ж, як величина самого хижака. Виявилося, що бероє харчуються виключно іншими реброплава. Таким чином, вони не тільки не є конкурентами планктоноядних риб, але, навпаки, їх слід вважати корисними тваринами, так як вони інтенсивно винищують дійсно шкідливих болінопсісов.
З'ясувалося й інше не менш важлива обставина: бероє служать їжею таким промисловим рибам, як пікша й тріска; поїдають їх і інші риби, їдять реброплавів великі сцифоїдних медузи.
Таким чином, розкривається одна із сторін найскладніших взаємовідносин між морськими тваринами. Коли до берегів підходить велика кількість тріски, вона виїдає майже всіх берое. Форма тіла реброплавів в значній мірі залежить від їхнього способу життя і тому дуже різноманітна. Найбільш типовою є овальна, яйцевидна або сигарообразная форма тіла, характерна для всіх реброплавів, забезпечених щупальцями, і для личинок багатьох видів, які в дорослому стані щупалець позбавлені. Вище вже йшлося про реброплава болінопсісе, який не має щупалець, але забезпечений двома великими ротовими лопатями. Не має щупалець і дорослий гребневик венерин пояс (Cestus veneris), що мешкає в Середземному морі. Тіло цієї тварини має стрічкоподібними форму і може сягати півтора метрів довжини. Венерін пояс плаває за допомогою гребних пластинок і внаслідок змієподібних рухів тіла, розташованого завжди в горизонтальній площині. Він прозорий і переливає всіма кольорами веселки, завдяки чому йому і дано таке поетичну назву.
Дуже своєрідні платіктеніди-маленькі сплощені гребневики, здатні не тільки плавати в товщі води, але й повзати по дну. До цієї групи відносяться описана А. О. Ковалевським целоплана Мечникова (Coeloplana metschnicowi) і описана Коротневим ктеноплана Ковалевського (Ctenoplana kowalewskii), а також ще близько двох десятків видів. У будові цих реброплавів можна угледіти дуже багато рис будови, загальних з деякими морськими плоскими червямі.Висказивалось навіть припущення, що плоскі черв'яки походять від плазунів реброплавів. Проте останнім часом завдяки дослідженням проф. В. М. Беклемішева стало ясно, що схожість між повзаючими реброплава і плоскими черв'яками чисто зовнішнє, залежне від однакового способу життя цих тварин, а також від схожості механізму їх ембріонального розвитку.
Один арктичний вид реброплавів - тьяльфіелла (Tjalfiella) є сидячим донним тваринам. Молодь цього виду веде планктонний спосіб життя, але потім сідає на дно ротом вниз, причому середина рота заростає, а його краї у вигляді двох трубок загинаються догори. До бесщупальцевим реброплава відносяться також згаданий вище бероє та інші близькі до нього види. Тіло бесщупальцевих реброплавів має вигляд мішка. Воно забезпечене величезним ротовим отвором, що служить для заковтування великої здобичі-реброплавів інших видів.


Тип Реброплави включає єдиний клас з шістьма загонами. Представники перших п'яти загонів (підклас щупальцевих - Tentaculifera) - цідіпповие (Cydippea), лопастеносние (Lobata), цестіди (Cestidea), платіктеніди (Platictenidea) і тьяльфелліди (Tjalfiellidea) - протягом всього життя або на ранніх стадіях розвитку мають щупальця. До підкласу бесщупальцевих (Atentaculata) відноситься лише один загін морських огірків (Beroidea), що включає реброплавів, позбавлених щупалець на всіх стадіях їх розвитку.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ6. Загальна характеристика Типу Плоскі черви (Plathelminthes)
Питання:
1. Загальна характеристика типу. Особливості зовнішньої та внутрішньої будови.
2. Клас війчасті черви розповсюдження, значення; Клас Сисуни - паразити людини та тварини.
3. Стьожкові черви паразити людини та тварини. Заходи боротьби з ними.
Тип налічує понад 12 тис. видів, серед яких є як вільноживучі, так і паразитичні форми. Еволюційні особливості плоских червів: виникнення мезодерми — джерела формування нових органів; подальше диференціювання клітин, поява м'язової тканини, представленої окремими м'язовими волокнами; білатеральна (двобічна) симетрія тіла; формування систем органів: нервової, з концентрацією нервових клітин біля переднього краю тіла; травної, що включає передній і середній відділи; видільної, представленої протонефридіями; статевою, що складається із статевих залоз та їхніх придатків.
Тіло плоских червів являє собою шкірно-м'язовий мішок, що складається з шару епітеліальних клітин (покривний епітелій) і трьох шарів м'язів — кільцевого (зовнішнього), діагонального (проміжного) та поздовжнього (внутрішнього). Простір між органами заповнений особливою тканиною — паренхімою, утвореною з клітин з відростками.
Нервова система складається з нервового вузла на передній частині тіла та поздовжніх нервових стовбурів, що відходять від нього. Останні сполучені між собою поперечними перетинками. У багатьох видів є примітивні органи чуттів — очі та статоцисти.
Видільна система представлена протонефридіями — системою каналів, які пронизують усе тіло тварини та відкриваються назовні порами. Кінцевий відділ протонефридія утворений особливою клітиною з безліччю війок, ритмічний рух якої створює потік рідини каналами до пор.
Дихальна і кровоносна системи відсутні. Поглинання кисню і виділення вуглекислого газу відбувається по всій поверхні тіла. У плоских червів рівномірний розподіл доживних речовин по тілу шляхом дифузії неможливий, тому відсутність кровоносної системи компенсується безліччю розгалужених виростів кишки.
Травна і статева системи у різних представників мають украй різноманітну будову. Характерною особливістю всіх плоских червів є гермафродитизм.
Тип Плоскі черви включає класи: Війчасті черви, Сисуни, Стьожкові черви. Останні два класи представлені виключно паразитичними формами.
Клас Війчасті черви (Turbellaria). Переважна більшість війчастих червів — вільноживучі види. Типовий представник класу — молочно-біла планарія, хижак, мешканець прісних вод (рис.12).
У покривному епітелії планарій є два види клітин: війчасті і залозисті, які секретують слиз. Завдяки узгодженому рухові війок і слизовому чохлу планарії швидко пересуваються в товщі води. Слиз виконує також захисну функцію. Під епітелієм розташовані три шари м'язових волокон. Планарія живиться дрібними молюсками, членистоногими.Ротовий отвір розміщений на черевному боці і веде до глотки, яка продовжується розгалуженим, сліпо замкненим кишечником. У просвіт кишки виділяються травні ферменти, проте велику роль виконує внутрішньоклітинне травлення. Неперетравлені залишки викидаються через рот. Розчинені продукти обміну виводяться каналами протонефридіїв. Нервова система представлена нервовим вузлом і нервовими стовбурами. Вони мають примітивні очі.

Турбелярії — гермафродити. Кожна особина має чоловічі (сім'яники) і жіночі (яєчники) статеві залози. Перші продукують сперматозоїди, а другі — яйцеклітини.Сперматозоїди однієї особини виходять крізь спеціальні вивідні протоки і за допомогою копулятивного органу потрапляють у жіночі статеві шляхиіншої особини, дезапліднюють яйцеклітину. Деякі турбелярії здатні до нестатевого (вегетативного) розмноження, при якому тіло тварини розділяється на дві частини.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 7. Особливості організації представників Класу Дигенетичні сисуни Diginea, Trematoda.
Питання:
1.Риси пристосування до паратизму, гетерогенез.
2.Життєві цикли та спосіб життя окремих представників (сисун печінковий – Fasciola hepatica, сисун котячий – Opistorchis, сисун ланцетоподібний – Dicrocoelum dendriticum, двоустка кров'яна – Schistosoma haematobium), та їх практичне значення.
3.Клас Моногететичні сисуни –Monogenoidea. Особливості будови і розвитку як ектопаразитів риб (дактилогірус – Dactylogyrus vastator, гіродактилус – Gyrodactilus, двiйчак парадоксальний – Diplozoon paradoxum. перехід від ектопразитизму до ендо-паразитизму – багатоуст жаб’ячий (Polystomum integerrimum).
Зберігаючи всі основні ознаки типу Плоскі черви, сисуни характеризуються специфічними особливостями, зумовленими паразитичним способом життя:
наявність спеціальних органів прикріплення (присосків, гачків);
тенденція до спрощення травної системи і здатність всмоктувати поживні речовини крізь покриви тіла;
відсутність війчастого епітелію;
високий рівень розвитку статевої системи та складні життєві цикли із зміною хазяїв.
Зовнішня будова (форма тіла) сисунів сплющена, листоподібна. Є два присоски — передній (ротовий), у глибині якого розташований рот, і задній (черевний). Покриви не несуть війок, шари м'язів добре розвинені.
Нервова система представлена нервовим ганглієм і поздовжніми нервовими стовбурами. Органи чуттів спрощені у зв'язку з паразитичним способом життя.
Рот веде до м'язистої глотки, яка виконує роль насоса під час ковтання їжі. Кишечник закінчується сліпо, але має у великих форм безліч виростів. У багатьох сисунів, добре пристосованих до паразитизму, травна система спрощується. Ці види здатні всмоктувати поживні речовини всією поверхнею тіла.

Більшість сисунів — гермафродити. Чоловіча статева система представлена двома сім'яниками, сім'явивідними протоками і ко- пулятивним органом. Жіноча статева система складається з яєч- ника і складної системи каналів, якими виводиться яйцеклітина. Незважаючи на гермафродитизм, запліднення у більшості сисунів перехресне: чоловічі гамети однієї особини запліднюють яйцеклітину іншої, і навпаки.
Печінковий сисун паразитує в жовчних протоках печінки великої рогатої худоби і людини.
У циклі розвитку печінкового сисуна спостерігається зміна хазяїв. В остаточному хазяїні відбувається статеве розмноження, у проміжному — партеногенетичне (рис.13).
Котячий сисун має схожий життєвий цикл; у нього два проміжні хазяї. Перший — молюск. Личинки, що залишають молюска, прикріплюються до другого проміжного хазяїна — риб (плотва, лящ). Поїдаючи рибу, кішки, собаки й інші рибоїдні ссавці заражуються паразитом. Людина може заразитися унаслідок вживання мороженої або недостатньо просоленої риби.
Особливістю паразитичних тварин є виняткова плодючість. Кожна особина може відкладати до декількох мільйонів яєць за день. Це пов'язано з дуже малою вірогідністю зустріти проміжного й остаточного хазяїна. Більшість яєць і личинок гинуть під дією не сприятливих чинників середовища.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 8. Загальна характеристика Класу Стьожкові черви (Cestoda)
Питання:
1.Риси пристосування до паразитизму в кишечнику хазяїна: розчленування тіла, органи прикріплення, відсутність травної системи.
2.Життєві цикли стьожка широкого (Diphillobotrium latum), ціп'яка свинячого (Taenia solium), ціп'яка бичачого (Taeniarhynchus saginatus), карликового ціп'яка (Humenolepis nana), альвеококу (Alveococus sp), ехінококу (Echinococus granulosus).
3.Практичне значення плоских червів та заходи боротьби з ними. Філогенія плоских червів.
Тіло стьожкових червів стрічкоподібне, поділене на членики — від декількох члеників в ехінокока до декількох тисяч у бичачого ціп'яка. На передньому кінці тіла знаходиться голівка (сколекс), забезпечена органами прикріплення присосками та гачечками. На голівці є безліч чутливих волосків — органів дотику.
Стьожкові черви повністю втратили травну систему. Вони всмоктують поживні речовини поверхнею тіла. Покриви тіла мають безліч виростів — ворсинок, що значно збільшує їхню поверхню та полегшує поглинання поживних речовин.
У кожному членику в більшості видів знаходяться чоловічі та жіночі статеві органи. Запліднення часто перехресне, але може бути і самозапліднення. У життєвому циклі відбувається зміна хазяїв.
Серед небезпечних паразитів людини треба виділити стьожака широкого та ехінокока. Зараження людини стьожаком проходе після вживання не провареної чи мало просмаженої риби. Тривалий термін просолювання риби може знищити личинок паразитів.
Остаточним хазяїном ехінококу і різні м’ясоїдні тварини (собаки, кішки, вовки, лисиці), проміжним – різні домашні травоїдні тварини та людина. Пильне ставлення до домашніх тварин, їх дегельменталізація може зменшити ризик захворювання паразитами.
Бичачий, або неозброєний, ціп’як (Taeniarhynchus saginatus) у статевозрілому стані паразитує в людини, проміжним хазяїном служить велика рогата худоба (звідси назва «бичачий»). Паразит розповсюджений майже у всіх країнах земної кулі.
Доросла цестода досягає довжини 4— 10 м; це один із самих довгих стрічкових хробаків. Ширина кінцевих члеників- 5-7 мм. Голівка, як й у всіх ціп’яків, несе чотири присоски, однак вона не має добре розвиненого хоботка й позбавлена гаків (звідси назва «неозброєний»). Стробила складається більш ніж з тисячі члеників. Особливості будови ціп'яків тісно пов'язані з необхідністю пристосування до паразитичного способу життя. Травна система у них повністю відсутня. Живляться паразити поживними речовинами, які знаходяться в кишках, всмоктуючи їх всією поверхнею тіла. Тіло їх вкрите типовим шкірно-м'язовим мішком, але покриви (тегумент) здатні одночасно протидіяти перетравлюючій дії травних соків і всмоктувати продукти із порожнини кишок людини. Органи чуттів відсутні. Пристосуванням до паразитичного способу життя є добре розвинена статева система і гермафродитизм, а також надзвичайна плодючість (у кожному членику бичачого ціп'яка знаходиться до 175 тис. яєць, за добу із організму їх виводиться близько 5 млн). Членики здатні самостійно виповзати через анальний отвір, а також, вийшовши з фекальними масами назовні, розповзатися по землі і по траві, збільшуючи тим самим можливість зараження. Велика рогата худоба заражається при ковтанні члеників або яєць.
Життя бичачого ціп’яка в кишечнику людини докладно вивчена радянськими вченими Ж. К. Штромом і Ф. Ф. Тализіним. Ці вчені проковтнули зрілі личинки ціп’яка, заразили себе цим паразитом і протягом декількох років вели ретельні спостереження за станом свого організму й життям розвинених у них ціп’яків.
Перебуваючи в кишечнику людини, ціп’як періодично відокремлює від тіла зрілі членики. У члеників при цьому відкривається кінець центрального каналу, і яйця можуть виходити через отвір, що утворився, назовні. Більша частина члеників виділяється назовні з випорожненнями, але деяка частина може виходити активно, тому що членик, що відірвався, має здатність до повільного пересування. Самостійне виходження спостерігається звичайно вночі під час сну хворого, але іноді це може відбуватися й удень.
Членики, що покинули кишечник, можуть плазувати по тілу й по білизні. За добу хворої виділяє до 28 члеників, якщо врахувати активно вийшли й з випорожненнями. У цих члениках з організму людини щодоби виділяється близько 5 млн. яєць ціп’яка.
У кишечнику людини бичачий ціп’як може жити більше десяти років.
Дорослий паразит у стьожковій стадії живе в тонкій кишці людини, а личинки його живуть в різних органах великої рогатої худоби. Хазяїна, в тілі якого розвиваються личинкові стадії, прийнято називати проміжним, з в тілі якого паразит досягає статевої зрілості — остаточним. Отже, для ціп'яка неозброєного проміжний хазяїн — велика рогата худоба, а основний — людина.
Велика рогата худоба — проміжний хазяїн бичачого ціп’яка — заражається паразитом, заковтуючи яйця його разом із забрудненою травою. У зрілому яйці втримується шестикрючная личинка – онкосфера. У кишечнику тварини онкосфера звільняється від оболонок і проникає в кровоносні судини кишечнику. Із кров’ю вона попадає в різні органи й у тканинах їх перетворюється в наступну стадію личинки – у фіну (цистицерк), що являє собою пухирець розміром приблизно з горошину, усередині якого перебуває сколекс цестоди, укручений у шийку на зразок пальця рукавички.
Найчастіше цистицерки, інакше вони називаються фіни, локалізуються в мускулах. Однак вони можуть перебувати під шкірою, в очах, у різних залозах, у мозку й інших органах.
Людина заражається бичачим ціп’яком, споживаючи в їжу сиру або погано проварену яловичину, уражену цистицерками.
У людей, заражених бичачим ціп’яком, спостерігається нудота, блювота, болю в животі, нестійкий стілець. Хворі люди страждають підвищеною дратівливістю, безсонням, запамороченням, іноді припадками, схожими з епілептичними. Відомі випадки закупорки кишечнику клубками паразита й проникнення їхніх тіл у жовчний міхур і підшлункову залозу. Особливо важко переносять захворювання діти.
Поразка цистицерками бичачого ціп’яка великої рогатої худоби проходить звичайно непомітно. Але при сильному зараженні, коли поряд з м’язами дивується мозок і серце тварин, хвороба проявляється різко й тварини, особливо молодняк, іноді гинуть.
Боротьба з тениаринхозом (захворювання, викликуване дорослою формою бичачого ціп’яка) проводиться шляхом виявлення й лікування хворих людей, а також проведення загальних санітарно-побутових заходів. Велике значення в боротьбі із цим захворюванням має виявлення й вибракування заражених цистицерками туш великої рогатої худоби, що вбиває на бойні. Цієї ж міри сприяють запобіганню тварин від зараження личиночними формами паразита. З метою скорішого викорінення цієї хвороби необхідно виявляти і лікувати хворих людей з обов'язковим знезаражуванням фекалій, не використовувати м'ясо, яке не пройшло ветеринарно-санітарного контролю, піддавати термічній обробці використовуване в їжу м'ясо і проводити санітарно-просвітницьку роботу серед населення.
Свинячий, або збройний, ціп’як (Taenia solium) відрізняється від бичачого меншими розмірами тіла (довжина його 2—3 м) і будовою сколекса. У цього виду ціп’яка сколекс має добре розвитий хоботок, що збройний двома рядами гаків (звідси назва ціп’яка- «збройний»). Є відмінності й у будові полової системи: двулопастной яєчник постачений маленькими додатковими часточками, матка з меншим числом бічних відростків – їх усього 7-12. Оторвавшиеся від тіла, зрілі членики свинячого ціп’яка не здатні до самостійних пересувань.
У дорослій стадії свинячий ціп’як паразитує в кишечнику людини. Проміжними хазяями його є домашні й дикі свині (звідси назва- «свинячий»).
Свині заражаються личинками ціп’яка при поїданні корму, у який попадають яйця паразита. Особливо часто заражаються свині в невпорядкованих господарствах, де вони нерідко харчуються нечистотами, що містять іноді не тільки окремі яйця, але й цілі членики цестоди.
Личинки свинячого ціп’яка, називані також цистицерками або фінами, улаштовані по такому ж типі, як й у бичачого, локалізуються вони головним чином у м’язах свиней. Крім м’язів, личинки можуть вражати й інші органи.
Людина заражається свинячим ціп’яком при вживанні в їжу незнешкодженого свинячого м’яса, що містить цистицерки паразита. При захворюванні проявляються симптоми, подібні з тими, які описані для хворих, заражених бичачим ціп’яком. Однак для людини свинячий ціп’як становить більшу небезпеку, чим бичачий. Справа в тому, що для свинячого ціп’яка людин може бути не тільки остаточним, але й проміжним хазяїном. Потрапивши тим або іншим шляхом в організм людини, яйце свинячого ціп’яка здатно розвиватися тут у цистицерка, викликаючи захворювання, називане цистицеркозом. Характер захворювання цистицеркозом людини залежить від того, у якому органі й у якій кількості перебувають личинки паразита. Якщо вони локалізуються в підшкірній клітковині, у м’язовій сполучній тканині або в м’язах, важкого захворювання може не бути. Але цистицерки можуть проникати в мозок або в очі. Захворювання в цих випадках супроводжується сильними головними болями, блювотою, припадками, подібними епілептичним, і іншими нервовими явищами, ведучими нерідко до смертельного результату. Поразка очей викликає часткову або повну втрату зору.
Видалення цистицерков з мозку або очей можливо тільки хірургічним шляхом.
Боротьба з тениидозом людини й цистицеркозом свиней проводиться в такий же спосіб, як і із захворюваннями, викликуваними дорослої й личиночной формами бичачого ціп’яка. Про їх ми говорили вище.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
Тип Круглі черви або Нематоди (Nemathelminthes)
Питання:
Особливості будови круглих червей.
Небезпечні паразити людини.
 Профілактика
нематодозів.
Профілактика
нематодозів.
Тип налічує близько 20 тис. видів, вільноживучих і паразитичних, що включають морські, прісноводні та ґрунтові форми. Еволюційні особливості
будови:
втрата паренхіми й поява первинної порожнини тіла — щілини між внутрішніми органами і стінкою тіла, заповненою рідиною;
диференціювання м'язової тканини на окремі тяжі поздовжніх м'язів;
поява задньої кишки, що відкривається назовні задньопрохідним (анальним) отвором.
Тіло нематод кругле в поперечному зрізі. Стінка тіла складається з еластичної та щільної кутикули, яка виділяється клітинами епітелію і м'язів. Уздовж тіла тягнуться чотири поздовжні м'язові тяжі (рис.14).
Нервова система розвинена сильніше, ніж у плоских червів. Спостерігається концентрація нервових вузлів біля переднього кінця тіла й утворення так званого навкологлоткового нервового кільця. Від нього відходять поздовжні нервові стовбури, сполучені перетинками. У багатьох нематод розвинені органи дотику, хімічного чуття.
Травна система наскрізна і ділиться на три відділи: передню, задню і середню кишку. У деяких паразитичних нематод розвивається позакишкове травлення, за якого ферменти виводяться в зовнішнє середовище, а напіврідка харчова кашка поглинається за допомогою спеціально пристосованого ротового апарату.
Система виділення нематод двох типів: канали виділень, що тягнуться уздовж всього тіла, і особливі клітини, що виділяють розчинні продукти обміну на поверхню тіла.
Круглі черви роздільностатеві. Самці мають сім'яники, сім'явивідні протоки і копулятивний орган; самки — яєчники і яйцепроводи. Чоловічі гамети не мають джгутиків, пересуваються амебоїдними рухами, тому одержали назву спермії.
Людська аскарида паразитує в тонкому кишечнику людини. Довжина тіла самок досягає 40см, самців — 25см. Запліднені яйця виводяться назовні з калом — для їхнього розвитку обов'язково потрібен кисень. На повітрі розвивається личинка, яка для подальшого розвитку повинна потрапити в організм людини. Зараження
відбувається через їжу, на яку яйця паразита можуть бути занесені мухами, тарганами. У тонкому кишечнику людини личинки звільняються від оболонки яйця, проникають у кровоносну систему, по ній укапіляри легень, далі в альвеоли, бронхи, глотку. Потім повторно зтуютьсяізновупотрапляютьу тонкий кишечник, де перетворюються на дорослих аскарид. Таким чином, у життєвому циклі аскариди, як і більшості нематод, є тільки один хазяїн (рис.15).
Гострик паразитує в прямій кишці людини, головним чином у дітей.
Він живиться вмістом кишки й бактеріальною флорою. Для дозрівання запліднених яєць необхідний кисень, тому самка ночами виповзає крізь анальний отвір назовні, відкладає яйця, що викликає свербіж у ділянці промежини. Яйця з личинками, що розвиваються в них, залишаються під нігтями дитини і легко потрапляють до рота, досягають товстого кишечника і перетворюються на дорослі особини (рис.16).
З апліднена
самка тримає в собі близько 12000 прозорих
яєць. Всередині яйця знаходиться звернута
личинка, вкрита зверху двошаровою
оболонкою. Самки після відкладки яєць
самки гинуть, але у дівчат та у жінок
вони існують досить тривалий термін, -
заповзають через статеву щілину до
піхви. При цьому гострики сприяють
захворюванню жіночої статевої системи
трихомонадами. Яйця гостриків достигають
на поверхні тіла людини на предметах
вжитку. Особливо часто хворіють
ентеробіозом повторно діти, тому, що
мають звичай ссати пальці та плохо мити
руки. Тому проведення суспільної
профілактики та підтримання гігієнічних
норм може стримати ріст захворення.
апліднена
самка тримає в собі близько 12000 прозорих
яєць. Всередині яйця знаходиться звернута
личинка, вкрита зверху двошаровою
оболонкою. Самки після відкладки яєць
самки гинуть, але у дівчат та у жінок
вони існують досить тривалий термін, -
заповзають через статеву щілину до
піхви. При цьому гострики сприяють
захворюванню жіночої статевої системи
трихомонадами. Яйця гостриків достигають
на поверхні тіла людини на предметах
вжитку. Особливо часто хворіють
ентеробіозом повторно діти, тому, що
мають звичай ссати пальці та плохо мити
руки. Тому проведення суспільної
профілактики та підтримання гігієнічних
норм може стримати ріст захворення.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
5. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.-
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.
Загальна характеристика Типу Коловертки (Rotatoria).
Питання:
1.Особливості будови. Розмноження і розвиток.
2.Зміна партенонентичних і статевих поколінь. Цикломорфоз. 3.Значення в живленні жаб.
Тип Скреблянки (Acantocephala).
1.Загальна характеристика.
2.Пристосування до паразитизму в кишечнику (органи прикріплення, редукція травної системи) інших тварин.
3.Розмноження та розвиток. Представники: скреблянки осeтрова (Acantocephalus), скреблянка велетень (Macracanthorhynchus hirudinaceus). Практичне значення.
Коловертками називають своєрідну групу в основному прісноводних мікроскопічних тварин, виділених в додатковий клас типу Nemathelminthes. Коловертки - самі дрібні з багатоклітинних тварин (від 0, 04 до 2 мм). За своїм систематичного становищем вони стоять серед нижчих сколецід (нижчих черв'яків - турбеллярий, нематод, гастротріх). Разом з тим вони вражають спостерігача своєю красою, дивовижною різноманітністю і тонкощами організації та біології. Вражає виняткова розмаїтість їх рухів (плавання, повзання, «крокування», нерухоме існування) і пов'язаних з ними будови покривів тіла і «будиночків» (у сидячих форм), а також війкового апарату.
Різноманіття форм руху пов'язано у коловерток з надзвичайною спеціалізацією війкового озброєння і розвитком поперечносмугастої мускулатури, що складається з окремих тяжів. Не менш притаманна для коловерток різноманітність пристосувань до зміни кліматичних умов, що проявляється в явищах сезонного цикломорфозу, в різноманітті статевих циклів і в здатності переходити в стан спокою.
Всіма цими особливостями коловертки різко виділяються не тільки серед нижчих безхребетних, а й взагалі серед всіх тварин.
Найбільш типовими утвореннями, за якими одразу можна відрізнити коловерток від інших тварин, є навколоротового апарат і жувальна ковтка, так званий «мастаки». Навколоротового апарат являє собою сукупність різному розташованих на передньому кінці тіла війок, які у деяких видів розташовуються по краях дисковидні виростів голови і своїм биттям, що нагадує мерехтіння спиць швидко обертового колеса, викликають коловороті, за що вони отримали свою назву. Коловерток називають також «ротатор», від латинських слів rota - колесо і fero-носити (rotiphera - колесонесущіе). Цей ресничний апарат служить для пересування і загарбання їжі.
Мастаке - розширена передня частина травної трубки (глотки) зі спеціальною мускулатурою, з залозами і нервами. Усередині мастаке знаходиться жувальний апарат з твердих частин, званих щелепами. У процесі еволюції у коловерток виробилися найрізноманітніші пристосування до різних умов існування. Така пластичність організації сприяла тому, що коловертки населяють не тільки різноманітні прісні, солоноватоводние і солоні басейни як з дуже високою, так і з дуже низькою температурою води, але багато хто з них пристосувалися до існування в місцях, де вода буває або в незначній кількості, або тільки тимчасово: у прибережному піску водойм, у моху, в лісовій підстилці, в дуплах дерев, у грунті. Основна маса видів коловерток населяє прісні водойми, в морях коловерток мало, але все ж таки є що живуть тільки в море. Крім видів своєрідного загону Seisonidea, є виключно морські представники родів: Synchaeta, Trichocerca, Lindia, Proales, Keratella та ін Значно більше видів зустрічається в опріснених частинах морів; це ті форми, які потрапляють у море з річок і добре пристосовуються до підвищеної солоності, причому деякі види в нових умовах навіть інтенсивніше розвиваються, ніж у прісній воді (Asplanchna brightwellii, Filinia longiseta, F. brachiata). Небагато коловертки ведуть паразитичний спосіб життя. Так, в черв'яках паразитують Drilophaga (рис. 245, 12), Albertia і Balatro; в відкладених яйцях прудовіков зустрічається Proales gigantaea, в соняшник - Proales latrunculus; є паразити водоростей, наприклад: у Vaucheria живе Proales wernecki, в колоніях Volvox - Proales parasita . У хребетних тварин і у людини коловертки не паразитують. Деякі види коловерток співмешкають з іншими організмами. Так, на дафнія живуть Proales daphnicola (рис. 245, 2), харчуючись водоростями, що обростають панцир рачка; Brachionus rubens (рис. 245, 3) часто і надовго прикріплюється ногою теж до панциру дафній, де, не витрачаючи енергії на пересування, в спокійному стані відфільтровує харчові частинки, весь час змінюючи за допомогою рачка місце облову.
Переважна більшість видів коловерток космополіти, вони зустрічаються на всій земній кулі, але є і властиві тільки Америці, наприклад Brachionus gessneri (рис. 245, 5), Br. dolabratus (рис. 245, 6), Br. havanaensis та ін, або тільки Старому Світу, як Brachionus diversicornis (рис. 245, 4) у Br. forficula; в Байкалі живуть види, що зустрічаються тільки в цьому озері, наприклад Notholca baicalensis, N. olehonensis (рис. 245, 7), Synchaeta pachipoda (рис. 245, 8) та ін; в Антарктиді знайдені коловертки, ніде більше не зустрінуті. Деякі звичайні види коловерток живуть у найрізноманітніших водоймах, вони невибагливі до умов існування - була б вода і їжа, їм не важливо, солона або прісна вода, проточна або стояча, тепла або холодна, вони майже при всіх умовах добре себе почувають і розмножуються. Такі організми називаються убіквістамі, тобто зустрічаються скрізь, усюди. Ши рокое поширення багатьох видів пов'язано з легкістю їх перенесення на великі відстані в сухому вигляді: у піявковідних коловерток - дорослих самок, а в інших - лежать яєць. Але чимало коловерток, які пред'являють суворі вимоги до умов існування, до якості їжі, до хімічного складу води і її фізичним показникам, і далеко не в кожній водоймі бувають потрібні їм умови. Якщо потрапила в даний водойму Коловертки не знаходить у ньому необхідних їй умов, вона просто гине. Такі види зустрічаються не часто і відносяться до «рідкісним». Таким чином, поширення коловерток по земній кулі в більшості випадків проблема не стільки зоогеографічна, скільки екологічна, так само як розподіл їх по водоймах різних типів та їх окремих місцях (прибережні зарості, дно, товща води).
Природно, що коловерток стали вивчати з часу винаходу мікроскопа, тобто з кінця XVII ст. Натуралісти XVIII і початку XIX ст. об'єднували коловерток з найпростішими, і тільки в 1817 р. Кюв'є вперше виділив коловерток у самостійну групу тварин, об'єднаних їм у розділ Radiata.
Статева система самця складається з великої семенника - статевої залози, що виробляє чоловічі статеві клітини - сперматозоїди, додаткових (передміхурова) залозок, вивідного протоку і копулятивного органу. У більшості коловерток самці народжуються вже з цілком розвиненими сперматозоїдами. Сперматозоїди відносно великі, складаються з голівки і вібруючого хвостика; їх небагато, найбільше число (близько 300) відомо у Asplanchna. У багатьох видів семенник підвішений на широкій стрічці, яка є рудиментом травного каналу. У самців деяких видів планктонних коловерток на спинній стороні, приблизно в середині тіла, лежить велика жирова капля, яка перешкоджає обертальному руху самця навколо осі свого тіла. Самці плавають прямо і в порівнянні з самками дуже швидко. Самці живуть не більше двох діб, адже вони нічого не їдять, витрачаючи за своє коротке життя тільки ті поживні речовини, які їм дісталися від матері. Найбільш активними вони бувають у перші години після вилуплення. Процес копуляції, тобто передачі сперматозоїдів самці, у деяких видів відбувається через клоаку, в інших - через будь-яке місце тіла самки. Статева система самки, що відноситься до загону Monogononta, складається з яєчника, жовточників і яйцепровода, що відкривається в клоаку.

Видільна система представлена типовими протонефридіями .У порожнині тіла з боків проходять канали, що починаються в голові і відкриваються у пульсуючий сечовий міхур, що є у деяких видів розширену клоаку, в інших - розширені кінці каналів. Вміст міхура виділяється в клоаку.
Нервова система коловерток складається з мозку, що лежить над глоткою в головному відділі (мозковий ганглій), гангліїв мастакс і ноги, дрібних гангліїв, розкиданих по багатьох місцях тіла, і нервів, що відходять від мозку, що пов'язують ганглії і йдуть до органів почуттів і м'язовим тяжам, Органи почуттів представлені очима і щупальцями. Очі - червоні або чорні пігментні плями. У деяких видів перед плямою є лінза (світлозаломлюючу тільце). У більшості коловерток одне «потиличне» око, що лежить поблизу мозку; у багатьох по два ока з боків голови; є «лобові» очі; в деяких піявковідних коловерток очі перебувають на хоботку; частина видів коловерток зовсім позбавлена очей, а у прикріплених форм очі мають тільки молоді особини. Щупальця представлені особливими клітинами з чутливими щетинками, віями або сосочками. Вони численні в навколоротового апараті; на спинній стороні знаходиться одне дорзально щупальце, у деяких видів дуже довге; з боків тіла лежать латеральні щупальця; є щупальця в порожнині глотки і ножні щупальця. Щупальця виконують функції органу дотику, статичного почуття і сприйняття хімічних подразнень.
Нервова система коловерток складається з мозку, що лежить над глоткою в головному відділі (мозковий ганглій), гангліїв мастакс і ноги, дрібних гангліїв, розкиданих по багатьох місцях тіла, і нервів, що відходять від мозку, що пов'язують ганглії і йдуть до органів почуттів і м'язовим тяжам, Органи почуттів представлені очима і щупальцями. Очі - червоні або чорні пігментні плями. У деяких видів перед плямою є лінза (світлозаломлюючу тільце). Над мозком у багатьох видів коловерток, але далеко не у всіх, є орган невідомого значення, званий ретро-церебральним, тобто органом «навпаки мозку». Він складається з досить великого мішка, часто заповненого чорними великими зернами, і що лежать по боках його двох субцеребральних залоз. У деяких видів є вивідні протоки цього органу, що відкриваються на тильній стороні голови. Цей загадковий орган за походженням належить до шкірних залоз.
Коловертки харчуються водоростями, бактеріями і детритом, крім того, є хижаки, що харчуються тваринами - найпростішими і коловертками, які значно менше хижака, але хижаки можуть харчуватися і водоростями. Переважна більшість коловерток полифаги, тобто мало обмежені у своєму виборі їжі, але є і монофаги, тобто такі, які харчуються тільки одним родом їжі, наприклад детритом або тільки певним видом водорості.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 9. Тип Кільчасті черви (Annellida) Клас Багатощетинкові (Poliychaeta)
Питання:
2. Особливості зовнішньої та внутрішньої будови.
3. Значення полі хет в природі і житті людини.
Описано близько 8 тис. видів кільчастих червів, що відзначаються різноманітністю внутрішньої організації. У типі виділяють три класи: Багатощетинкові, Малощетинкові, П'явки.
Еволюційні особливості будови:
поява вторинної порожнини тіла (целому), яка відрізняється від первинної тим, що має власну стінку, утворену епітелієм;
виникнення кровоносної системи;
організація нервової системи за типом черевного нервового лан- цюжка;
поява нового типу системи виділення — метанефрйдіальної;
розчленовування тіла на сегменти (метамерія).
Сегментація тіла — основна ознака кільчастих червів. Метамерія виникає в процесі еволюції при подовженні тіла. Число сегментів різне — від 5—6 до 500—600.
У кільчастих червів формуються параподії — органи руху у вигляді виростів тіла кожного сегмента, з яких назовні направлені війки. Параподії добре розвинені у багатощетинкових, а у малощетинкових від них залишаються лише щетини; п'явки позбавлені навіть щетинок.
У багатьох представників типу (дощового черв'яка) дихальна система відсутня. Проте деякі багатощетинкові мають зябра — вирости шкірного покриву, розташовані на голові або параподіях.
Дощовий черв'як — характерний представник класу Малощетинкові.
У тілі дощового черв'яка виділяють голову і тулуб, який поділений на окремі кільця — сегменти. Кожен сегмент (за винятком першого) несе чотири пучки війок—органів руху. Стінка тіла складається з декількох шарів:
1) тонкої неклітинної кутикули, яка виділяється шкірою;
2) шкірного епітелію з одного шару клітин;
3) двох шарів мускулатури — зовнішнього кільцевого і внутрішнього поздовжнього;
4) епітелію, що обмежує целом.
Целом заповнений рідиною і розділений тонкими перегородками на окремі відсіки, які відповідають зовнішній сегментації.
Нервова система характеризується подальшою концентрацією нервових клітин біля переднього кінця тіла. Надглотковий нервовий вузол великий (його іноді називають головним мозком). Нижче розташований підглотковий вузол.Вузли сполучені двома тяжами, що охоплюють глотку з утворенням навкологлоткового нервового кільця. На черевному боці тіла утворюється єдиний нервовий стовбур — черевний нервовий ланцюжок, що починається від підглоткового нервового вузла. У кожному сегменті він утворює нервовий вузол, від якого відходять нерви. З органів чуття розвинений дотик.
Травний канал проходить уздовж усього тіла від ротового отвору до анального. Він підрозділяється на відділи: рот, глотку, стравохід, зоб, шлунок, середню і задню кишку.
Кровоносна система замкненого типу. Кров тече по спинній і черевній судинах, від яких у кожному сегменті відходять кільцеві судини меншого діаметру до внутрішніх органів і м'язів. У шкірі утворюється густа мережа капілярів. Серця немає; його роль виконує спинна судина, стінки якої здатні до ритмічних скорочень
Дихання здійснюється всією поверхнею тіла. Кисень дифундує в капіляри шкіри і доставляється до органів.
Видільна система складається з метанефридіїв — по два в кож

ному сегменті, Метанефридій являє собою канал, що відкривається лійкою в целом, а протилежним боком — назовні. Він відрізняється від протонефридію багатоклітинністю.
Кільчасті черви — гермафродити. Чоловіча статева система представлена сім'яниками і сім'явивідними протоками (10—11-й сегменти), жіноча — яєчниками і яйцепроводами (13-й сегмент). Запліднення перехресне. На поясочку — залозистому потовщенні шкіри певних сегментів, виділяється слизовий кокон, що має форму барильця. Він містить яйцеклітини, які під час проходження кокона крізь сегменти з сїм'яприймачами запліднюються сперматозоїдами. У дощового черв'яка розвиток прямий.
Багатощетинкові здатні до нестатевого (вегетативного) розмноження. Тіло черв'яка розпадається на декілька фрагментів, у кожного з яких розвиваються ті частини тіла, яких бракує. Це свідчить про високу здатність до регенерації.
Величезна кількість видів (в основному багатощетинкові) мешкає в морях і океанах, як тропічних, так і помірних, навіть полярних широт, у прибережних районах або біля дна; деякі пристосувалися до існування в солонуватих водоймищах. Малощетинкові нерідко зустрічаються в прісних водах (річках,озерах,болотах), проте більша частина видів заселила ґрунт. Дощові черв’яки харчуються головним чином рослинними залишками, наявними в грунті, а також листям і стеблинками трав, які вони утягують в свої нірки. З таким способом живлення і пов'язане заковтування черв'яком землі. Органи травлення черв’яка мають набагато складнішу будову в порівнянні з кишковою порожниною гідри. Усередині тіла черв’яка є особлива порожнина, що відокремлена від кишкової порожнини. У цій порожнині тіла і розташований кишечник і інші внутрішні органи. Порожнина тіла розділена поперечними перетинками, відповідними по положенню зовнішніх члеників черв'яка. Кишечник і інші органи проходять крізь ці перегородки. Таким чином, дощовий черв’як має членисту будову не тільки зовні, але і всередині.
Кишечник черв’яка має складну будову, що пов'язано із способом живлення. Він складається з декількох відділів стравоходу, зобу, шлунку і кишки. Остаточна зміна і всмоктування перевареної їжі відбувається в кишці. Земля з неперетравленими оостаками їжі віддаляється з кишечника через заднепроходное отвір. Це звичайно і відбувається ночами, коли черв'яки виповзають на поверхню грунту.
Завдяки своєму способу життя і способу живлення дощові черв’яки мають дуже велике значення в процесі утворення грунту. Разом з іншими грунтовими тваринами дощові черв'яки постійно розпушують грунт і цим полегшують доступ в неї повітря і вологи. Вони перемішують грунт, оскільки виносять землю, що проковтнула ними, з глибших шарів на поверхню. Цим черв'яки як би перепахують грунт. Але особливо важливо, що, пропускаючи грунт через свій кишечник, харчуючись рослинними залишками, вони значно прискорюють процес утворення перегною. Підраховано, що за рік дощові черв'яки виносять на поверхню одного гектара до 20т і більше землі, що пройшла через їх кишечник. Особливо велике значення робота дощових черв'яків має для молодих посадок лісу.
Це найчисленніший клас анелід (понад 5000 видів, близько
130 видів живе у водоймах України). Здебільшого поліхети — жителі
морських водойм (населяють весь Світовий океан) і лише зрідка —
солонуватих і прісних водойм. Окремі представники ведуть паразитичний
спосіб життя. І Найхарактернішою ознакою багатощетинкових є На параподіях у багатьох видів поліхет містяться розгалужені зябра. Всі
поліхети роздільностатеві, розвиток із перетворенням; личинка — вкрита
війками трохофора веде планктонний спосіб життя. Тривалість життя
поліхет —2—3 роки. Багато поліхет гине " під час розмноження,
оскільки виведення статевих продуктів у них пов'язане із розривами
стінок тіла.
Цей клас поділяють на два підкласи: Рухливі і Сидячі. Перші за допомогою
параподій активно переміщуються по дну, плавають, енергійно проникають у
грунт. У них добре розвинений головний відділ з очами та іншими органами
чуття. За способом живлення вони найчастіше хижаки.
Сидячі форми живуть переважно в трубках, утворених із речовин, що
виділяються їхньою шкірою, та піщинками, уламками черепашок, грудочками
мулу. Іноді ці трубочки майже повністю занурені в грунт, з якого
вистиркається лише передня частина тіла з численними щупальцеподібними
відростками, що утворюють ловильний апарат. Головний відділ розвинений
слабко або редукований. Параподії також розвинені слабко.
До підкласу Рухливих належать нереїди, які зустрічаються у всіх морях
Радянського Союзу, в тому числі в Чорному та Азовському.
У 1939—1941 pp. під керівництвом академіка Л. О. Зенкевича (1889—1970)
були проведені роботи по переселенню нереїд із Азовського моря в
Каспійське. Акліматизація пройшла успішно і сприяла зростанню харчових
ресурсів Каспію. Живлячись детритом (рослинними рештками), нереїди
включають у харчові ланцюги мул, який раніше «мертвим капіталом» лежав
на дні Каспійського моря. Самі ж вони мають високу калорійність (1 г
сухої речовини містить до 5000 кал) і становлять основну кормову, базу
для осетрових риб. Осетер і севрюга майже повністю живляться нереїсом.
До підкласу Сидячих належать піскожили (рис. 35), що зустрічаються в
північних і далекосхідних морях, а також у Чорному морі. Усі види
піскожилів живуть на літоралі, найчастіше на піщаних мілинах,
зариваючись у грунт на глибині до 20—30 см. Вони викопують дугоподібні
нірки з двома отворами на поверхні грунту. Піскожил захоплює і проковтує
пісок разом із органічними рештками, якими він живиться. Пісок проходить
крізь кишки і викидається ззаду. Тому біля рота піскожила на поверхні
грунту утворюється лійка, а позаду його тіла — конусоподібні купки
землі. У лійку потрапляють гниючі рослинні рештки, якими він живиться.
Коли піскожил викидає з нірки чергову порцію піску, його можуть схопити
риби. Проте жертвує він лише своїм хвостом, який через деякий час
регенерує.
Особливості організації Класу Малощетинкових червів (Oligochaeta)
1.Адаптація до риючого способу життя.
2.Значення олігохет у підвищенні родючості грунту.
Травна система починається ротовим отвором на передньому кінці тіла і закінчується відхідником на останньому членику. Кишка складається з трьох відділів: переднього (ектодермального), середнього (ентодермального) і заднього (ектодермального). Передня кишка у дощового черв'яка представлена кількома відділами: глоткою, стравоходом, волом і м'язовим шлунком. Починається передня кишка ротовим отвором, далі йде овальна мускулиста глотка. Глотка переходить у вужчий і довший стравохід, у стінках якого розміщені вапнякові залози. У деяких хижих кільчаків є хітинові щелепи, призначені для захоплення здобичі. На рівні 14—16го сегментів стравохід розширюється і утворює воло. З вола їжа переходить у так званий м'язовий шлунок, де й перетирається. Від шлунка майже до заднього кінця тіла тягнеться середня кишка, де під дією ферментів їжа перетравлюється і всмоктується. В стінці кишки з'являються шари м'язів, що забезпечують її самостійну перистальтику. Неперетравлені рештки переміщуються в коротку задню кишку і викидаються назовні через відхідник.
Дихальна система. У дощового черв'яка газообмін відбувається через багату на кровоносні судини шкіру; у деяких морських кільчаків є зябра.
Видільна система. В кожному членику дощового черв'яка є парний орган виділення, який складається з лійки і тоненьких трубочок. Продукти життєдіяльності з порожнини тіла потрапляють у лійку. Від лійки йде канадець, який переходить у сусідній сегмент, робить кілька петель і відкривається на бічній стінці тіла. Лійка і канадець мають війки, рух яких зумовлює переміщення виділюваної рідини. Такі органи виділення називають метанефридіями.
Кровоносна система. У більшості кільчастих червів вона замкнена. Уздовж усього тіла над органами травлення тягнеться спинна судина, а під ними — черевна. На передньому й задньому кінцях тіла вони сполучені між собою. В кожному сегменті є кільцева судина, яка сполучає спинну й черевну судини. У передній частині тіла є кілька товщих кільцевих судин, це так звані "серця". За рахунок ритмічних скорочень "сердець" і спинної кровоносної судини забезпечується рух крові по спинній судині від задньої частини тіла до передньої, по черевній судині — навпаки, а по кільцевих судинах — від спинної до черевної судини. Кров у дощового черв'яка червона, в ній є гемоцити з гемоглобіном.
Нервова система складніша, ніж у плоских і круглих червів. Навколо глотки є навкологлоткове кільце: надглотковий і підглотковий вузли та перемички, що їх сполучають. На черевному боці розміщені два нервових стовбури, які мають у кожному членику потовщення — ганглії, з'єднані між собою перемичками. У багатьох кільчаків відбувається зближення правого й лівого нервових стовбурів, внаслідок чого утворюється черевний нервовий ланцюжок.
З органів чуття у кільчастих червів є вусики, вічка, органи рівноваги, які частіше розміщені на головному сегменті.
Регенерація. Як і гідра та війчасті черви, кільчаки здатні до регенерації, тобто відновлення втрачених частин тіла. Якщо дощового черв'яка розрізати на дві частини, то у кожної з них відновляться втрачені органи.
Статева система складається з жіночих гонад (яєчників), що являють собою комплекс статевих клітин, оточених епітелієм, і чоловічих гонад (сім'яників), що лежать усередині містких мішків.
Дощові черви — гермафродити, однак серед кільчаків трапляються і роздільностатеві форми. На тілі дощового черв'яка є поясок, який виробляє слиз. Запліднення перехресне, відбувається після копуляції двох особин. Під час копуляції відбувається обмін сперматозоонами. Після Цього черв'яки хвилеподібними рухами виповзають із слизової муфти. Коли муфта проходить повз отвори жіночих і чоловічих статевих залоз, у неї відкладаються яйця (власні) і сперматозоони (партнера), відбувається запліднення. Муфта перетворюється на кокон, у якому й відбувається прямий (без метаморфозу) розвиток дощових черв'яків.
Розвиток. У дощового черв'яка розвиток прямий, однак у деяких кільчаків із заплідненого яйця утворюється личинка, тобто розвиток відбувається з перетворенням.
Отже, кільчасті черви характеризуються низкою прогресивних ознак, до яких належать поява сегментації, вторинної порожнини, кровоносної та дихальної систем, а також підвищення організації видільної і нервової систем.
Дощові черви живляться рослинними рештками, що містяться в грунті. Вони пропускають його через кишки, залишаючи на поверхні купки екскрементів, до складу яких входить грунт. Цим вони сприяють перемішуванню й розпушуванню грунту, а також збагаченню його на органічні речовини, поліпшенню водного й газового режимів грунту. Ще Ч. Дарвін відмічав корисний вплив кільчастих червів на родючість грунту.
Нині дощові черв'яки потребують охорони, їхня чисельність може скорочуватися внаслідок надмірного використання добрив та пестицидів. Дощовий черв'як занесений до Червоної книги України.
Тип П'явки (Hirudinei) - ряд класу кільчастих хробаків. Тіло подовжене абр овальне, більш-менш сплющене в спинно-черевному напрямку, ясно розділене на дрібні кільця, які в числі 3 - 5 відповідають одному сегменту тіла; в шкірі численні залози, які виділяють слиз; на задньому кінці тіла зазвичай велика присоска, на передньому кінці є теж добре розвинена присоска, у центрі якої міститься рот, що служить для присмоктування. На передньому кінці тіла 1 - 5 пар очей, розташовані дугою або попарно один за одним. Порошиця на спинному боці над задньою присоскою.
Нервова система складається із дволопатевого надглоткового ганглію або головного мозку, з'єднаного з ним короткими комісурами під глоткового вузла і самого черевного ланцюжка, що міститься у черевному відділі. Кровоносній синус має близько 20 узлов.Головной вузол інервує органи чуття і глотку, а від кожного вузла черевного ланцюжка відходить 2 пари нервів, що інервують відповідні їм сегменти тіла; нижня стінка кишечника забезпечена особливим пповздовжнім нервом, що дає гілки до сліпого мішка кишечника.
Органи травлення починаются ротом, озбройним чотирьма хітиновими зубчастими пластинками (щелепні П. - Gnathobdellidae), що служать для прорізання шкіри при смоктанні крові у тварин, рот здатний випинатися хоботком (у хоботних П. - Rhynchobdellidae); в порожнину рота відкриваются багаточисельні слинні залози, що іноді виділяють отруйний секрет ; за глоткою, що грає при ссанні роль насоса, розташований великий еластичний шлунок, забезпечений бічними мішками (до 11 пар), з яких задні найдовші; задня кишка тонка і коротка.
Кровоносна сістема сскладаєтьсячастиною зі справжніх, пульсуючих, судин, частиною з порожнин-синусів, що представляють залишок порожнини (вторинної) тіла і з’єднанихних між собою кільцевими каналами; кров у хоботних П. безбарвна, у щелепних - червона, внаслідок розчиненого в лімфі гемоглобіна.
Особливі органи дихання є тільки у р. Branchellion, у формі листовидних придатків з боків тіла.
Видільні органи влаштовані по типу метанефридій або сегментованих органів кільчастих червів і у большінства П. їх є по парі у кожному з середніх сегментів тіла.
П. - гермафродити: Чоловічі статеві органи складаються у більшості з пухірців (сім'яників), по парі в 6 - 12 середніх сегментах тіла, з'єднаних на кожній стороні тіла загальною вивідною протокою; ці протокі відкриваются назовні одним отвором, розташованим на черевному боці одного із передніх кілець тіла; жіночий статевий отвір лежить на один сегмент позаду чоловічого і веде в два окремих яйцеводи з мішковидними яєчниками. Копулюють дві особини, кожна одночасно граючи роль самки ісамца. П. під час кладки яєць виділяє залозами, що лежать в області статевих органів, густий слиз, у вигляді чохла . П., в цей чохол відкладаються яйця, після чого П. виповзає ізнього, при цьому краї його отворів зближуються, склеюються і утворюють таким чином капсулу з яйцями всередині, прикріплену до поверхні листка водорості.
Всі П. - хижаки, що харчуються кров'ю здебільшого теплокровних тварин, або молюсків, черв'яків і т. п.; живуть вони переважно в прісних водоймах або у вологій траві, але є і морські форми (Pontobdella), як і наземні форми (у Цейлоні ). Hirudomedicinalis - медична П. до 10 см. у довжину 2 см. завширшки, чорно-бура, чорно-зелена, з повздовжнім візерунчастим червонуватим малюнком на спині; черево світло-сіре, з 5 парами очей на 3, 5 і 8 кільцях і сильними щелепами; поширена в болотах півд. Європи, півд. Росії і Кавказу. У Мексиці в медицині викорстовується Haementaria officinalis; інший вид, П. mexicana - отруйна; в тропічній Азії поширена у вологих лісах й у траві Hirudo ceylonica та інші види, які заподіюють хворобливі укуси людині і тваринам. Aulostomum gulo - кінська П., чорно-зеленого кольору, з більш світлим низом, має озброєння роту слабкіше і тому непридатна для терапевтичних цілей; самий звичайний вид у східній Європі. Nephelis vulgaris-невелика П. з тонким вузьким тілом, сірого кольору, іноді з бурим малюнком на спині; з 8-ма очима, розташованими дугою на головному конці тіла; споріднена їй оригінальна Archaeobdella Esmonti, рожевого кольору, без задньої присоски; живе на муловому дні в Каспійському і Азовському морях.Clepsiue tesselata - татарська П., з широкоовальним тілом, зеленовато-бурого кольору, з кількома рядами бородавок на спині і 6-ма парами трикутних очей, розташованих оди за одним , мешкає на Кавказі і в Криму, використовується татарами для лікувальних цілей; перехідне місце в ряді щетинконогих (Chaetopoda Oligochaeta) червів занімає Acanthobdella peledina, що зустрічається в Онезькому озері у Росії. П. медична (Hirudo officinalis) - водиться у нас П.висмоктує від 8 до 15 гр. крові. В лікувальних цілях їх приставляють від 2 до 12 і навіть до 25штук; вони повинні бути свіжі і здорові, швидко скорочуватися під час дотику. Так як перетравлення і всмоктування крові триває звичайно 5 - 9, іноді 12 - 18 місяців, то раніше спожиті П. рідко використовуються для нового використання. Потрібно вибирати П. середньої величини, вагою в 1 - 2гр., У віці 3 - 6 років. Ніколи не слід відривати їх насильно, так як в рані можуть залишитися щелепи П. Звичайно, насмоктавшись, вони самі відпадають; при необхідності - відпадання може бути прискорене присипкою на їх тіло солі або змазуванням його оцтом.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 10. Тип Молюски (Mollusca)
Питання:
1. Основні властивості м’якунів
2. Будова черепашки її значення.
3. Прогресивний розвиток внутрішніх органів.
4. Головоногі м’якуни – як вища ступінь еволюційного розвитку.
Тип включає понад 130 тис. видів, згрупованих у класи: Черевоногі, Двостулкові, Головоногі. Багато даних указує на спорідненість молюсків із стародавніми кільчастими або плоскими червами. Еволюційні особливості будови: поява дихальної системи; поява серця.
Тіло молюсків складається з голови, тулуба та ноги. Нога являє собою виріст черевної стінки тулуба і служить для пересування, ловлі здобичі, прикріплення, закопування в ґрунт. Характерна особливість молюсків — наявність черепашки. Із внутрішнього боку до черепашки прилягає шкірна складка — мантія. Простір між мантією і тілом називається мантійною порожниною. У порожнині знаходяться зябра, в неї відкриваються отвори органів виділення та анальний отвір.
Вторинна порожнина (целом) у більшості молюсків редукується і зберігається, у дорослих форм лише у двох ділянках — перикардїі та порожнині. статевої залози.
Нервова система складається із сукупності нервових вузлів, розташованих в ділянці голови, у нозі та деяких ділянках тіла. Вузли зв'язані між собою нервовими стовбурами. Органи чуттів розвинені неоднаково у різних видів. Деякі молюски мають очі (наприклад, виноградний слимак), в інших основну роль виконують органи хімічного чуття, рівноваги.
Органи дихання у морських видів представлені зябрами, а у наземних і прісноводних — легенями (виноградний слимак, ставковик). Легені розташовані в мантійній порожнині й оповиті густою мережею капілярів.
Травна система починається ротовим отвором, який веде до м'язистої глотки. Для ротового апарату характерна на'явність особливого органу — тертки (радули), забезпеченої хітиновими зубчиками. З її допомогою молюски зіскоблюють водорості з каміння, захоплюють здобич. У ротову порожнину відкриваються слинні залози. Далі йде стравохід, шлунок, у який відкривається протока травної залози («печінки»). Із шлунка бере початок кишечник, часто звитий. Анальний отвір відкривається в мантійну порожнину.
Кровоносна система незамкнена. У деяких ділянках тіла кров тече не по кровоносних судинах, а по порожнинах між внутрішніми органами. Серце складається зі шлуночка й одного або декількох передсердь. Серце розташоване в навколосерцевій сумці — перикардії, заповненій рідиною. У багатьох видів кров безбарвна, оскільки не має дихальних пігментів. У цьому випадку її називають гемолімфою. Гемолімфа виконує в основному транспортну функцію — переносить кисень і поживні речовини.
Видільна система представлена однією, рідше декількома нирками. Нирка — це канал, який одним кінцем сполучається з порожниною перикарду, а іншим відкривається в мантійну порожнину.
Багато молюсків — гермафродити, хоча деякі примітивні види роздільностатеві. У своєму розвитку молюски часто проходять стадію личинки, яка дуже схожа на таку у багатощетинкових червів. Ця стадія рухома і служить для розселення виду.
Клас Двостулкові (Bivalvia).
Клас включає 20 тис. видів, більшість з яких мешкають у морях, і лише деякі заселили прісні води (рис.20).
Черепашка двостулкових складається з двох стулок, сполучених еластичною зв'язкою. Кожна стулка вкрита зсередини мантією. У разі небезпеки молюск закриває черепашку. Це здійснюється завдяки роботі м'язів замикачів. Для підвищення сили скріплення стулок, на кожній з них часто є безліч зубчиків — замок. Голова і пов'язані з нею органи чуттів зредуковані. Відсутня також глотка, слинні залози, тертка.

У мантійній порожнині двостулкових розташовані зяброві листки, які постійно омиваються потоком води. Вода надходить у порожнину крізь трубкоподібний виріст мантії — дихальний сифон, а видаляється крізь вивідний сифон.
Циркуляція води всередині мантійної порожнини забезпечує молюсків не тільки киснем, але й їжею. Зважені у воді частинки (органічні залишки, планктон) завдяки роботі особливих війкових клітин прямують до ротового отвору і заковтуються. Непридатні для їжі частинки видаляються крізь вивідний сифон. Двостулкові профільтровують значні об'єми води — 5—10 л за годину (устриці, мідії).
Органами дотику служать чутливі клітини, розсіяні в епітелії І зокрема груповані в навколоротових щупальцях, по краю ноги, по краю мантії тощо. Біля основи зябер е пара осфрадіїв, будова яких нагадує будову осфрадіїв у примітивних черевоногих. Близько біля педальних гангліїв розміщена пара статоцистів, які мають таку саму будову, як і в інших молюсків. У зв'язку з редукцією голови головних щупальців і очей немає, проте нерідкі випадки вторинного виникнення органів зору по краю мантії на щупальцях, що оточують отвори сифонів тощо. Такі вторинні очі можуть мати різну будову. Найбільш прості з них являють собою очні ямки, вистелені епітеліальними та пігментними клітинами. Проте у деяких форм вторинні очі мають більш складну будову і належать до типу пухировидних. Оригінальної будови очі розміщені по краю мантії у гребінців. Вони являють собою випинання з віялоподібним розміщенням клітин, до складу яких входять зорові клітини конічної форми, циліндричної форми, пігментні і так звані інтерстиціальні клітини.
Розмноження і розвиток. Переважна більшість пластинчатозябрових роздільностатеві. Статева система їх складається з гонад і вивідних проток. Залози є продуктом целома. Вони парні, розміщені в передньому відділі тулуба і заходять в основу ноги. Лише в деяких випадках гермафродитні форми мають дві пари залоз; у більшості ж гермафродитів з кожного боку розміщено по одній гермафродитній залозі з однією вивідною протокою, хоча у деяких є цілком відокремлені частини цієї залози, що продукують лише сперматозоїди або лише яйцеклітини. Статеві клітини формуються з зачаткового епітелію, яким встелені гонади. У більшості випадків статеві продукти виводяться назовні через протоки, що відкриваються по боках ноги в мантійну порожнину поруч з отворами нефридіїв, але у більш примітивних форм статеві продукти виводяться через нефридії.
Копулятивного апарата немає. Більшість пластинчатозябрових відкладає яйця у воду, де й відбувається запліднення. Найчастіше яйця відкладаються по одному, і лише зрідка спостерігається прикріплення яєць до підводних предметів. Проте у багатьох прісноводних форм яйця відкладаються в зябра, де і відбувається розвиток до виходу личинки. Так, у процесі розвитку морських форм утворюються личинки трохофорного типу. Внаслідок поступових змін трохофора перетворюється в типову для пластинчатозябрових личинкову форму — велігер (парусник). На цій стадії личинка значно наближається до дорослої форми, відрізняючись від неї наявністю паруса. Перетворення парусника на дорослу форму.

На відміну від морських, у прісноводих двостулкових розвиток відбувається абсолютно своєрідно. Так, у беззубки з яєць, що виношуються в зябрах, виходять особливі личинки - глохідії (рис.21).
Глохидії мають двостулкову раковину трикутної форми, з гострими зубцями посередині краю кожної стулки. Між стулками раковинки глохідія—сильний мускул-замикач. Крім того, глохидий має бисусну залозу, що виділяє бисусную нитку. У беззубки глохідії розвиваються восени і зимують в зябрах матері. Весною вони викидаються у воду і прикріпляються до шкіри, зябер і плавників риб за допомогою клейкої биссусной нитки і зубців раковини. Потім, під впливом роздратування шкіри риби, починається обростання глохідія епітелієм шкіри господаря, і утворюється бульбашка у вигляді маленького нариву з глохідієм всередині.
У такому стані глохідій паразитує на шкірі риби протягом двох і більше місяців. Потім шкірна бульбашка лопається і що розвинувся до цього часу з глохідія молодий молюск падає на дно. Такий своєрідний спосіб розвитку забезпечує розселення молюсків за допомогою риб. У інших прісноводих двостулкових, наприклад, у кульовок зародки розвиваються в мантийной порожнині в особливих «виводкових камерах» на зябрах, З мантийной порожнини виходять крихітні молюски, що вже цілком сформувалися.
Клас Черевоногі, або Слимаки (Gastropda).
Клас включає близько 85 тис. видів, розміром від декількох міліметрів до 7—15 см. У більшості видів видів черепашка закручена в спіраль і вкриває все тіло (рис.18). У слимаків вона маленька і частково або повністю накрита виступаючою мантією. У деяких видів черепашка повністю редукується.
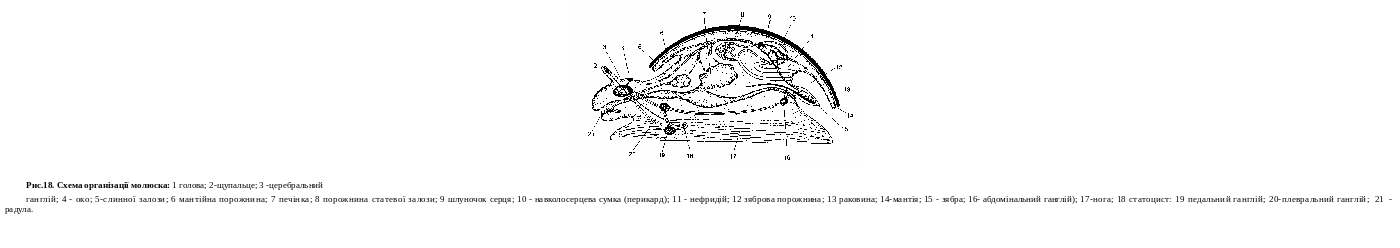
Нога, як правило, розвинена добре і служить органом пересування. У багатьох слимаків задня частина ноги має рогову кришечку, яка щільно закриває устя, коли молюск ховається в черепашку.
Серед черевоногих є як роздільностатеві, так і види гермафродитів. Запліднення в більшості випадків внутрішнє. Розвиток може проходити з утворенням личинки або без неї.
Черевоногі пристосувалися до надзвичайно різноманітних умов існування. Вони заселили прибережні зони океанів, морів та їхні глибини (рапани), прісноводні водоймища (ставковики), частина видів пристосувалася до життя на суші (виноградний слимак, слимаки), заселивши навіть пустелі та вершини гір. У наземних форм захистом від несприятливих умов середовища (посухи, холоду) служить затягування отвору черепашки плівкою слизу, який твердне на повітрі. Органами дихання прісноводних і наземних видів є легені.

Наземні молюски мають на голові дві пари щупалець (органи дотику); на кінцях однієї з них розташовані очі (Садовий слимак) (рис.19). У прісноводних форм очі знаходяться біля основи єдиної пари щупалець, їжа черевоногих надзвичайно різноманітна: планктон, водні та наземні рослини, черви, ракоподібні, інші молюски.
Клас Головоногі. Описано понад 700 видів головоногих. Головоногі мають рудиментарну черепашку. Ротовий отвір оточений щупальцями (вісім у восьминогів, десять у каракатиць і кальмарів). На щупальцях у декілька рядів розташовані присоски. Голова велика (рис.22).
Головний мозок головоногих вкритий зачатковою корою, зверху захищений хрящовим черепом. Орган зору — очі, акомодація досягається видаленням або наближенням кришталика до сітківки. Смакові рецептори розташовані на язиці та присосках щупалець. Органи рівноваги представлені статоцистами.
Кров має блакитний колір, що пов'язано з присутністю пігменту, який містить купрум — гемоціаніну, який бере участь у транспорті кисню. У головоногих три серця: головне складається з передсердя і шлуночка й забезпечує потік крові по тілу,а інші два (зяброві серця) проштовхують її крізь зябра.
Рот невеликий, м'язиста глотка має роговий дзьоб. Головоногі - хижаки. Вони харчуються рибою, крабами, черепашками, нерідко зустрічається канібалізм.

Головоногі роздільностатеві. Запліднення внутрішнє: самець щупальцем переносить сперматозоїди в статевий отвір самки. Розвиток прямий, без личинкової стадії.
Рух здійснюється за принципом реактивного двигуна. Крізь мантійний отвір вода засмоктується в мантійну порожнину, а потім з силою виштовхується крізь вивідний сифон. При цьому молюск пересувається заднім кінцем тіла вперед. Деякі восьминоги можуть «ходити» по дну, спираючись на щупальця.
Особливості організації типу Оніхофор (Onychophora)
Питання:
1.Риси будови, що зближують їх з кільчастими червами та членистоногими.
Єдиний клас - первиннотрахейні(Protracheata), розділений на дві родини: Peripatidae - мешкають в тропічних районах Мексики, Центральної і півночі Південної Америки, екваторіальної Західній Африці і на південно-сході Азії.
Peripatopsidae - Чилі, Південна Африка, Австралія і Нова Зеландія.
У Північній півкулі зустрічаються лише в Мексиці. Приблизно 90-110 сучасних видів; водяться у вологих місцях, головним чином у тропіках. Всі сучасні види - наземні тварини.
У викопному вигляді відомі з кембрію (Aysheaia, Xenusion - останнього іноді відносять до пізнього протерозою), причому як мешканці морів. Деякі вже нагадували сучасних онихофор; інші були озброєні шипами (Hallucigenia). Іноді до онихофор відносять також копалин лопастеногіх епохи кембрію і ордовика (типовий представник: аномалокаріс).
Тіло онихофор гусеніцеобразное, вкрите м'якою кутикулою з безліччю поперечних складок. Довжина тіла від 2 до 20 см (Macroperipatus, острів Тринідад); самці дрібніші за самок. Забарвлення найрізноманітніша, від яскравої (червоною, синьою) до тьмяною.
На голові - пара м'ясистих вусиків і пара очок. У ротовій порожнині є пара гачковидний щелеп. У самців деяких видів на голові також є відростки для передачі сперматофор. Тулуб з 14-43 парами нерозчленованих ніг - виростів стінки тіла, озброєних слабкими кігтиками; у самців ніг зазвичай менше. У самців на задніх ніжках є особливі залози, які виділяють феромон, який приваблює самок. М'язова система у вигляді шкірно-м'язового мішка, характерного для хробаків. М'язи зовнішнього шару кільцеві, внутрішнього - поздовжні. Травна система у вигляді трубки. Органи дихання - пучки трахей, дихальця яких розкидані по всій поверхні тіла. Видільні органи відкриваються у підстав ніг. Кровоносна система незамкнена - гемолімфа надходить в серці, трубчастий спинна судина з отворами, з порожнини тіла. Порожнина тіла при цьому функціонує як гідростатичний скелет; ноги також наповнені кров'ю і мають спеціальні регулюючі клапани. Нервова система сходового типу: від надглоткового вузла відходять два поздовжніх нервових стовбура, з'єднаних між собою перемичками-коміссур.
Через наявність спільних рис онихофор часто називають сполучною ланкою між кільчастими черв'яками та членистоногими. Як у кільчастих хробаків, у них сегментоване тіло з м'якою стінкою, нерозчленовані придатки, парні нефрідіі в кожному сегменті і неветвістий травний тракт. З членистоногими їх зближує трахейне дихання і редукція целома: простір між внутрішніми органами зайнято гемоцелем - порожниною, заповненою кров'ю.
Мешкають Оніхофори в тропічних і субтропічних лісах - у листовій підстилці, в гниючих пнях і під камінням; зустрічаються і на березі моря, в купах водоростей. Справа в тому, що тонка (в 1 мк) кутикула і відкриті отвори трахей майже не перешкоджають випаровуванню води, тому онихофор доводиться постійно жити у вологому середовищі.
Оніхофори - активні хижаки, що харчуються термітами, павуками та іншими дрібними безхребетними. На полювання виходять вночі або під час дощу. Спосіб полювання дуже своєрідний: щоб знерухомити видобуток, вони викидають клейку слиз із спеціальних залоз, розташованих з боків рота. У деяких видів «плювок» здатний вразити ціль на відстані до 30 см. Травлення у онихофор зовнішнє, як у павуків: їхній укус впорскується в тіло здобичі слину, що містить травні соки і ферменти; потім онихофор втягує вийшов «бульйон» ротом.
Роздільностатеві. Статева система самців, крім сім'яників і семяпроводів, що відкриваються в загальний канал, включає додаткові залози, які виробляють оболонку сперматофор - особливі мішечки, в які упакована сперма. Сперматофори, самці деяких видів (Peripatopsis) прикріплюють прямо до тіла самок, після чого в цьому місці кутикула розчиняється і сперматофор занурюється в порожнину тіла, сперматозоїди виходять з нього і мігрують до яєчники. Але частіше сперматофори відкладаються в субстрат, і самки захоплюють їх статевим отвором.
Запліднення відбувається в організмі самки. Самки зазвичай яйцеживородні (зародок розвивається за рахунок жовтка, але вилуплюється ще в статевих шляхах самки) або живородні. У живонароджених видів у матці утворюється плацента, через яку до зародку надходять поживні речовини; розвиток зародків може тривати більше року.

Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
ЛЕКЦІЯ 11. Тип Членистоногі (Arthropoda)
Питання:
1. Основны ознаки членистоногих.
2. Загальна характеристика підтипу зябродихаючих.
3.Особливості будови надкласу Ракоподібні (Crustacea)
На сьогодні описано понад 1 млн видів членистоногих, згрупованих у класи: Ракоподібні, Павукоподібні, Комахи.
Еволюційні особливості будови:
поява досконалих органів пересування — кінцівок;
виникнення поперечносмугастої мускулатури;
розвиток міцного зовнішнього скелета;
високий рівень розвитку дихальної системи.
У тілі членистоногих виділяють відділи: голова, на якій знаходяться органи чуттів і органи захоплення їжі, груди, на яких розташовані органи пересування (кінцівки, крила), черевце. У деяких членистоногих голова і груди утворюють єдиний відділ тіла — головогруди.
Усі відділи тіла складаються з сегментів, кількість яких є важливою систематичною ознакою. Кінцівки складаються з декількох члеників і утворюють систему важелів, які приводяться в рух роботою м'язів. У різних сегментах вони виконують рухову, захисну, статеву та дихальну функції. Деякі кінцівки на голові перетворюються на ротові придатки — органи захоплення, утримання і подрібнення здобичі.
2)
3}
Тіло вкрите твердою оболонкою — кутикулою, основним компонентом якої є полісахарид хітин. Кутикула виділяється залозистими клітинами покривного епітелію.Вона утворює так званий зовнішній скелет. Хітиновий панцир не розтяжний, і ріст організму можливий тільки в період линьки.
Зовнішній скелет виконує такі функції:
1) захисну, перешкоджаючи механічним пошкодженням;
2) служить місцем прикріплення м'язів;
3) запобігає випаровуванню води з поверхні тіла (ця властивість дозволила членистоногим освоїти сушу).
М'язова система членистоногих досягла великого розвитку, Поперечносмугасті м'язи дуже сильні й здатні скорочуватися з величезною швидкістю.
Нервова система організована за принципом черевного нервового ланцюжка. Кожний сегмент тіла несе нервовий вузол. Концентрація нервових вузлів у головному відділі призводить до утворення головного мозку, що досягає великих розмірів у вищих комах.
З органів чуттів найбільшого розвитку у багатьох членистоногих набули нюх і зір. Очі можуть бути прості або складні — фасеткові.
Дихальна система у первинноводних членистоногих (ракоподібні) представлена зябрами — виростами кінцівок. Деякі дрібні водні та наземні паразитичні членистоногі поглинають кисень усією поверхнею тіла. У великих наземних видів розвиваються особливі органи дихання — легені, які являють собою мішкоподібні випини кінцівок (павуки, скорпіони). Найбільш довершені органи дихання — трахеї — мережа тонких розгалужених трубок (комахи). Трахеї відкриваються на черевці маленькими отворами дихальцями. Найменші з трахей мають діаметр у декілька мікрометрів і підходять до кожної клітини тіла.
Система виділення водних членистоногих представлена метанефридіями.
Кровоносна система незамкнена. З появою трахей вона частково або повністю перестає викопувати функцію транспорту дихальних газів.
Більшість
членистоногих роздільностатеві, але є
також види гермафродити
(деякі ракоподібні). Запліднення може
бути зовнішнім
(у водних), внутрішнім і зовнішньо-внутрішнім
- самці виділяють
сперматозоїди у зовнішнє середовище,
а с амки
підбирають їх
своїми статевими отворами (більшість
кліщів). Більшість
видів відкладає яйця в зовнішнє
середовище. Дуже рідко
спостерігається живородіння (деякі
скорпіони, кліщі), іноді
— яйцеживородіння (тля). Якщо особина,
що вийшла з яйця, різко
відрізняється від дорослої, її називають
личинкою. Ріст більшості
членистоногих має ступінчастий
характер—розміри тіла збільшуються
стрибкоподібно, відразу після линьки.
амки
підбирають їх
своїми статевими отворами (більшість
кліщів). Більшість
видів відкладає яйця в зовнішнє
середовище. Дуже рідко
спостерігається живородіння (деякі
скорпіони, кліщі), іноді
— яйцеживородіння (тля). Якщо особина,
що вийшла з яйця, різко
відрізняється від дорослої, її називають
личинкою. Ріст більшості
членистоногих має ступінчастий
характер—розміри тіла збільшуються
стрибкоподібно, відразу після линьки.
Клас Ракоподібні. Клас включає 30—35 тис. видів. З них найвищу організацію мають представники ряду Десятиногі раки (річкові раки, краби, омари, лангусти, креветки).
Тіло річкового рака складається з голови, грудей, черевця, анальної (хвостової) лопаті (рис.24). Голова утворюється злиттям п'яти сегментів і, отже, несе п'ять пар придатків (видозмінених кінцівок): передні та задні антени (вусики), три пари щелеп. Обидві пари антен є органами нюху, дотику та хімічного чуття. На голові розташовані фасеткові очі. Груди складаються з восьми сегментів. Перші три несуть три пари кінцівок — ногощелеп; їх основна функція — подача їжі до ротового отвору. Інші п'ять сегментів утворюють п'ять нар ходильних кінцівок, які служатьдляпересування. Перша пара - клішні - наймасивніша й бере участь у захопленні жертви. На грудних кінцівках розташовуються органи дихання - зябра. Черевце також сегментоване. П'ять пар черевних кінцівок служать для утримання яєць, що розвиваються. Шоста пара разом з останнім сегментом черевця утворюють анальну лопать, щобере участь у пересуванні.
Кровоносна система незамкнена. Серце має форму п'ятикутника, розташоване на спинному боці тіла й оточене перикардом.
Травна система починається ротовим отвором, оточеним трьома парами щелеп. Рот веде до короткого стравоходу, який переходить в об'ємистий шлунок. У жувальному відділі шлунка відбувається перетирання їжі за допомогою хітинових зубців, у цідильному відділі — її фільтрація. Далі їжа потрапляє в кишечник, де вона перетравлюється (у просвіт кишки відкривається вивідна протока печінки) і всмоктується. Неперетравлені залишки видаляються крізь анальний отвір в ділянці анальної лопаті.
Органи виділення — парні зелені залози, подібні до метанефридій кільчастих червів і розташовані в передній частині тіла. Вивід ні протоки відкриваються біля основи антен.
Річкові раки роздільностатеві. Запліднені яйця утримуються черевними кінцівками самки. Розвиток прямий.
Ракоподібні заселили морські та прісні води всіх кліматичних зон. Багато з них пристосувалися до життя на суші
У прісноводних водоймищах численними є дрібні рачки дафнії — представники ряду Гіллястовусі. Вельми поширені в прісних водоймищах і циклопи - маленькі, до 2 мм рачки з ряду Веслоногі. Більшість циклопів хижаки, але є також рослиноїдні види. Багато ракоподібних пристосувалося до паразитичного способу життя, що завдають шкоди рибам, на зябрах яких вони живуть.
Наприклад - паразитичні веслоногі рачки під впливом паразитичного способу життя придбали характерні пристосування. Антеннули і антени їх різним чином пристосовані до прикріплення до органів господаря і висмоктування соків з його тканин (рис.26).

Нарешті, необхідно відзначити, що деякі ракоподібні (прісноводі циклопи і диаптомусы) є проміжними господарями в життєвому циклі паразитичних черв'яків (широкого стожака, ришти і інших) (рис. 25).
На відміну від нижчих вищі ракоподібні мають на черевці кінцівки. Представник вищих раків - японский краб - проміжний господар легеневого сисуна. Багато вищих ракоподібних їстівні, мають смачне, живильне, високоцінне м'ясо (раки, креветки, лангуста, омари, краби).
Краби іноді досягають крупних розмірів. Тіло гігантського японського краба з тарілку, а ноги завдовжки в 1м. Камчатський краб важить 7кг, тіло його досягає в поперечнику 12—25 см. Серед вищих ракоподібних є і сухопутні тварини, наприклад стонога, що живе в сирих приміщеннях. Вона має (на відміну від інших ракоподібних) органи повітряного дихання.
Характеристика класів Зябронгі раки (Branchiopoda), Щелепоногі(Maxillopoda), Черепашкові рачки (Ostracoda), Вищі раки (Malacostraca).
Зяброногі (Branchiopoda) ракоподібні вважаються самими примітивними представниками всього класу ракоподібних. Їх голова не зростається з грудними сегментами. Грудні ніжки листоподібні, слабо хітинізовані і служать одночасно для пересування, дихання та напрямки їжі до рота. При їх помахах створюються потоки води, що використовується і для руху, і для харчування, і для дихання.
Число грудних сегментів не постійне, будова статевої, нервової і кровоносної систем примітивніша, ніж у інших ракоподібних.
Зяброногі мешкають переважно у прісних водах, морські види нечисленні і мають прісноводне походження.
Всі зяброногі ведуть вільний спосіб життя, серед них немає ні паразитів, ні сидячих-прикріплених видів. Вони не виробили пристосувань для існування в підземних водах і на великих океанічних глибинах.
У цей підклас об'єднуються два ряди нині живучих ракоподібних-зяброногі (Anostraca) і листоногі (Phyllopoda).
Щелепоногі (Maxillopoda) - клас водяних ракоподібних (Crustacea). Характерні риси цієї групи - редукція числа сегментів черевця і відсутність на них кінцівок. Голова складена п'ятьма сегментами, груди - шістьма, черевце - чотирма. Найбільш відомі представники цієї групи - циклопи (Cyclopidae) і балянуси (Balanus). Більшість мають менший розмір, включаючи найдрібнішого в світі представника типу Членистоногі, Stygotantulus stocki, що має довжину 0,1 мм. Вірогідно, челюстеногіе не є монофілетичної групою. На думку багатьох авторів, це скоріше службовий таксон, в якому зібрані всі загони ракоподібних, які не можуть бути віднесені до інших класів.
|
|
|
Веслоногі раки (Copepoda) |
Calanoida Cyclopoida Gelyelloida Harpacticoida Misophrioida Monstrilloida Mormonilloida Platycopioida Poecilostomatoida Siphonostomatoida |
|
Thecostraca |
Cirripedia Facetotecta Ascothoracida |
|
Карпоїди (Branchiura) |
Arguloida |
|
П’ятивустки (Pentastomida) |
Cephalobaenida Porocephalida |
|
Mystacocarida |
Ctenocheilocaris Derocheilocaris |
|
 Calocalanus
pavo
(Calanoida: Calocalanidae)
Calocalanus
pavo
(Calanoida: Calocalanidae) Chthamalus
stellatus
(Sessilia: Chthamalidae)
Chthamalus
stellatus
(Sessilia: Chthamalidae) Argulus
(Argulidae)
Argulus
(Argulidae) Armillifer
armillatus
(Porocephalidae)
Armillifer
armillatus
(Porocephalidae)
ряд Черепашкові ракоподібні (Ostracoda)
Мешкають у дрібних калюжах, в ставках, у великих озерах, в підземних водах, на всіх глибинах Світового океану, як на дні, так і в товщі води, можна виявити цих непоказних дрібних тварин. Більше того, один вид ракушкових - Mesocypris terrestris - перейшов до сухопутному існуванню: він живе в лісовій підстилці вологих тропічних лісів Південної Африки. Відомо близько 2000 видів черепашкових.
Незважаючи на настільки різні умови існування і різний спосіб життя (серед Черепашкових є хижаки, є і грунтоїди, донні та планктонні форми, зариваються в грунт і плазують по рослинах і т. д.), організація черепашкових загалом досить одноманітна. Всі вони володіють товстої двостулковими раковиною, відповідної карапакс інших ракоподібних. Внутрішня її поверхня хітинізована, а зовнішня звичайно вапняна. Ліва та права стулки раковини з'єднані на спинній стороні еластичної зв'язкою. У багатьох видів є замок, що складається з виступів на одній стулці і відповідних їм заглиблень на інший. Приблизно в середині тіла розташований потужний мускул-замикач, прикріплений до правої та лівої стулок. При скороченні цього м'язу раковина щільно закривається.
Форма і будова черепашки залежать від способу життя. У рачків, які живуть на поверхні жорсткого грунту, раковина зазвичай товстостінна і забезпечена різними горбами і виростами; у видів, які порпаються в грунті, вона товстостінна, але гладка, а у видів, що повзають по рослинах і живуть на м'якому мулистому грунті, вона тонка і гладка . Особливо тонкою раковиною мають морські планктонні види, питома вага тіла яких наближається до питомої ваги води.
Голова часто хітинізована. На ній зазвичай поміщається непарний науплиальний очей, а у представників морського сімейства Cypridinidae є пара складних фасеткових очей, що досягають у глибоководного планктонного Gigantocypris дуже великих розмірів. Ділянки раковини, що знаходяться перед очима, прозорі, так що рачки бачать через раковину.
Всі п'ять пар придатків голови добре розвинені. Найбільш потужна з них - задня антена, яка служить основним органом пересування. Вона багата різними щетинками, що здобувають у деяких видів форму маленьких клешнів. У багатьох прісноводних черепашкових передня антена також довга, хоча і тонша від задньої. Вони плавають, змахуючи передніми антенами вперед і вгору, а задніми вперед і вниз, рухаючись при цьому по рівнодіючої, тобто прямо. У представників переважно морської родини Cytheridae на кінці скороченої зовнішньої гілки задніх антен відкриваються протоки парної павутинної залози. Вертикально опускаючись з якого-небудь підводного предмета, рачки виділяють, як павуки, павутинову нитку, а потім можуть нею користуватися при поверненні назад. Ця нитка дозволяє їм також утримуватися на грунті і так протистояти течії.
Жвали забезпечені сильно розвиненими щупиками, які нерідко допомагають руху рачків. Деякі Cypridinidae, наприклад Asterope, зариваються в грунт за допомогою щупиків жвал. Щелепи у різних груп черепашкових влаштовані різному. Звичайно зовнішня гілка однієї з них перетворена на широку платівку, помахи якої змінюють воду в порожнині раковини. Дихання здійснюється через стінку тіла, і тільки в Asterope на задньому відділі тіла є спеціальні вирости покривів, що функціонують як зябра. У більшості черепашкових щелепи забезпечені спеціалізованими щетинками й беруть участь у захопленні їжі, але у Cytheridae задня щелепу служить для пересування рачка і являє собою ходильні кінцівку. Перша, а у деяких родин (Cytheridae, Darwinulidae) перша і друга пара грудних ніг також ходильні, але у Сурridae і у всіх представників морського підряду Myodocopa задні грудні ніжки загнуті на спинний бік і пристосовані для очищення мантійної порожнини: вони видаляють потрапивші туди дрібні сторонні частинки. Є родини, зовсім позбавлені обох пар (Polycope) або задньої пари (Cytherella) грудних ніжок. Задня частина тіла в більшості випадків закінчується добре розвиненою Фуркою(лопаттю), що приймає участь в русі рачка. Ці рухи дуже різноманітні. Багато прісноводні Cypridae можуть і плавати і повзати. При повзанні вони чіпляються щетинками задніх антен за субстрат і відштовхуються від нього передніми грудними ногами і Фурко; передні антени при цьому протягнуті вперед і намацують шлях. Деякі види (Notodromas monacha, Heterocypris incongruens, Cyproides marginata) можуть пересуватися черевною стороною вгору, підвішуючись знизу до поверхневої плівки натягу, як це робить описаний вище гілчастовусий рачок Scapholeberis. Інші здатні зариватися в грунт на глибину до 7 см. Види, пристосовані до пересування по поверхні грунту і рослинам, часто відрізняються подовженими задніми антенами і передніми грудними ніжками. Морські планктонні Conchoecia і Gigantocypris плавають за допомогою своїх потужних задніх антен. Про спосіб пересування єдиного сухопутного представника ряду - Mesocypris terrestris - нічого не відомо, так як спостерігати його живим не вдалося. Судячи з того, що задні антени дуже великі і озброєні міцними когтевіднимі щетинками, можна припустити, що вони служать основним органом пересування.
Також різні і способи харчування. Багато черепашкових харчуються переважно водоростями, залишками рослин і тварин та органічною речовиною грунту (Cytheridae, Darwinulidae, Candonidae, Cypridopsis). Серед морських видів є хижаки. У кишечнику Cypridina castanea виявлені залишки мізиди і кіленогіх молюсків, в кишечнику С. погvegica - залишки поліхет. Планктонні черепашкові поїдають інших планктонних тварин, в першу чергу веслоногих, але іноді і щетинкощелпних і дрібних риб, а деякі види Conchoecia поряд з зоопланктоном використовують і фітопланктон. Деякі родини (морські Asterope, Cyclasterope, Cytherella, прісноводний Notodromas) - справжні фільтратори з відповідно облаштованими ротовими придатками. Asterope і Cyclasterope зариваються в грунт, виставляючи над ним кінці передніх антен, що утворюють отвір у грунті, через яке проходить службовець для фільтрації струм води.
Всі черепашкові роздільностатеві. За зовнішнім виглядом самці майже не відрізняються від самок. Втім, у багатьох прісноводних Cypridae самки розмножуються партеногенетично, а самці взагалі невідомі. В умовах акваріуму Негpetocypris reptans розводили протягом 30 років, причому весь цей час відбувалося партеногенетичний розмноження і самці жодного разу не з'явилися. Є види, які в північній частині області поширення розмножуються партеногенетично, а в південній зустрічаються і самці і самки і спостерігається процес запліднення.Статеві органи черепашкових влаштовані дуже складно. Однією з найбільш дивних особливостей цих рачків можна вважати величезні розміри їх сперматозоїдів, що перевершують за розміром сперматозоїди всіх інших тварин. Так, наприклад, у черепашкового рачка Pontocypris довжиною 0, 7 мм довжина сперматозоїда досягає 6 мм, тобто виявляється в 8 разів більше самої тварини. Довжина сперматозоїда людини (0, 062 мм) майже в 100 разів менше, ніж рачка.Вміщають такі гігантські сперматозоїди статеві шляхи самців і самок в сильно звитих каналах. Самці мають складний копулятивний апарат, за допомогою якого через щілину раковини вводять сперматозоїди в статевий отвір самки. Яйця запліднюються при виході із статевого отвору і спочатку виявляються в порожнині раковини, в її задній частині. Через деякий час самка відкладає яйця на камені, поверхня грунту або на рослини. Деякі Cytheridae і Darwinulidae носять яйця під раковиною до виходу з них личинок, а у Cypridinidae під раковиною повністю формуються молоді рачки, після чого мати видаляє їх назовні своїми грудними ногами. У решти черепашкових з яйця виходить науплиус своєрідного будови. Він забезпечений двостулковою черепашкою, коротшою і сильніше звужується до заднього кінця, ніж черепашка дорослих рачків. Його задні антени і жвали одноветвісти, причому останні закінчуються довгим кігтем. Науплиус може не тільки добре плавати, але й повзати за допомогою задніх антен і жвал. Личинка линяє 5-8 разів, поступово набуває відсутні кінцівки і перетворюється на дорослого рачка. У прісноводних видів тривалість личинкового розвитку коливається від 5 тижнів до 4 місяців. На відміну від інших ракоподібних дорослі черепашкові не линяють. Прісноводні види, науплії яких вилуплюються з перезимували яєць, живуть зазвичай не більше 4месяців, види родини Candonidae - 9 і більше місяців, а морський Philomedes globosus - не менше 2 1 / 2 років.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛКЦІЯ 12. Загальна характеристика типу хеліцерові (Chelicerata)
Питання:
1.Пристосування ло наземного способу життя.
2.Ееологічна радіація павукоподібних і кліщів.
3.Практичне значення.
Клас Павукоподібні (Arachnida). Клас об'єднує 60 тис. видів, згрупованих у декілька рядів, найчисленнішими з яких є Павуки (27 тис.) і Кліщі (25 тис.). Скорпіони (близько 700 видів) — найдавніший ряд серед наземних ленистоногих, що зберіг безліч примітивних рис властивих водним предкам. Сучасні павукоподібні представлені наземними формами, що походять від водних членистоногих, які спочатку заселяли моря.

Тіло Павукоподібних складається з головогрудей і черевця. Головогруди утворені щільною кутикулою і включають 6 сегментів. Кінцівки першого сегмента — хеліцери — є органами захоплення і подрібнення їжі (рис.27). У павуків на кінчиках хеліцер відкриваються протоки отруйних залоз. Кінцівки другого сегмента — педіпальпи — виконують функцію утримання здобичі, беруть участь у перенесенні сперми в статеві шляхи самки, служать органами дотику.

Кінцівки наступних чотирьох сегментів — ноги — беруть участь у пересуванні, плетенні павутини (павуки), викопуванні нірок, утриманні яєчного кокона, здобичі. Вони забезпечені нюховими та дотиковими рецепторами.
На відміну від ракоподібних обидві антени в павуків редукуються, фасеткові очі відсутні.
Черевце з'єднується з головогрудьми за допомогою сьомого сегмента, який не несе кінцівок. У багатьох кліщів головогруди й черевце зливаються в єдине ціле і втрачають сегментацію. Кінцівки черевця, що несуть зябра, перетворюються на легені. Кінцівки десятого й одинадцятого сегментів утворюють павутинні бородавки.
Нервова сиситема характеризується подальшою концентрацією нервових вузлів. У павуків у передньому відділі головогрудей виникає єдине скупчення нейронів — головогрудна нервова маса. З органів чуттів розвинений дотик (чутливі волоски на тілі та кінцівках) і нюх. Зір дуже слабкий — є декілька пар простих очок.
Більшість павукоподібних — хижаки, пристосовані до живлення напіврідкою їжею (рис.28). Спіймана здобич вбивається отрутою, покриви жертви розриваються і всередину вводяться травні соки. Потім розріджений вміст жертви всмоктується. Для цього служить забезпечена м'язами глотка, яка переходить у тонкий стравохід і далі — у сисний шлунок. Вирости кишечнику утворюють печінку, клітини якої виділяють травні ферменти.
Дихальна система представлена легенями або трахеями. У вищих павуків виникають трахеї. Деякі павуки мають і легені, і трахеї. Кліщі завдяки дрібним розмірам пристосувалися до поглинання кисню всією поверхнею тіла.
Кровоносна система добре розвинена у видів, які дихають легенями. На спинному боці головогрудей розташоване серце, від якого відходять великі кровоносні судини.
Система виділення представлена мальпігієвими судинами — сліпими виростами кишечнику.
Павукоподібні роздільностатеві. Запліднення в більшості випадків внутрішнє. Поширені повсюдно. Вирішальне значення в еволюції павуків відіграла павутина (тенета). Вона є притулком, пасткою, місцем спаровування, механізмом розповсюдження. Нитка павутини утворюється на повітрі з секрету павутинових залоз, розташованих у порожнині черевця. Павутинні залози відкриваються назовні павутинними бородавками.
У павуків виражений статевий диморфізм. Зазвичай самець менший за самку (іноді в тисячу разів), і часто після запліднення поїдається нею. Запліднення внутрішнє. Яйця відкладаються в кокон, зроблений з павутини. Багато видів піклуються про потомство — охороняють кокон, доглядають за ним.
Підтип трахейнодихаючі (Tracheata)
1.Загальна характеристика надкласу Багатоніжки (Myriapoda).
2.Спосіб життя та поділ на класи, їх характеристика.
Багатоні́жки (лат. Myriapoda) — надклас наземних трахейнодишних членистоногих тварин, які характеризуються нетагматизованим, гомономно сегментованим тілом. Лише передні членики тіла зливаються в головну капсулу — акрон. Кожен членик тіла несе пару або дві пари кінців Голова.Складається з акрона, у Симфіл голова вкрита справжньою головною капсулою, яка подібна до такої ж у комах. На голові містяться також очі: у Губоногих вони прості або складні; у Двопарноногих — представлені скупченням простих вічок; у Пауропод та Симфіл — взагалі відсутні. Перша пара тулубних кінцівок у Губоногих перетворилися на ногощелепи із серпоподібним кігтем, який містить отруйну залозу, в інших утворюються мандибули, а у Пауропод — ще й одна пара мандибул. Вусики прості або розгалужені (у Пауропод).Тіло.Тулуб гомономний, складається з різної кількості члеників, причому у більшості видів немає сталого їх числа. На кожному сегменті міститься пара кінцівок, у Двопарноногих — по дві пари, а у Пауропод 1-4 членики позбавлені ніг.
Нервова система. Типової для членистоногих будови: вона складається з головного мозку, навкологлоткових комісур, підглоткового ганглія та черевного нервового ланцюжка. У Двопарноногих подвоєні нервові ганглії черевного нервового ланцюжка в усіх сегментах, починаючи з V, а у Симфіл черевний ланцюжок має завжди 12 гангліїв.
Кровоносна система. Кровоносна система добре розвинена у Губоногих, а у Паропод взагалі відсутня. Серце, у вигляді довгої трубки, що лежить над кишечником уздовж тулуба. На задньому кінці воно сліпо замкнене. Серце поділене на камери, кожна з яких має два бічні отвори (остії). Спереду серце продовжується в головну аорту. По боках від аорти відходить артеріальне кільце, що огинає кишку і впадає в черевну судину (відсутня у Двопарноногих). Крім остій, що ведуть до навколосерцевої сумки, від кожної камери відходять по дві бічні артерії. У Симфіл кровоносна система дуже редукована. Кров впадає в лакуни порожнини тіла, з яких знову повертається в навколосерцеву сумку, звідки через остії потрапляє до серця. По серцевій судині кров рухається від заднього кінця до переднього, а по черевній — у зворотному напрямку.
Травна система. Для травної системи характерна наявність слинних залоз та відсутність у середній кишці печінкової, або травної залози.
Видільна система. Органи виділення представлені однією парою мапьпігієвих судин. У деяких Губоногих та Симфіл зберігається одна-дві пари максилярних залоз, як у Ракоподібних.
Дихальна система. Органи дихання представлені трахеями — тоненькими розгалуженими трубочками ектодермального походження, що виникають як глибокі занурення покривів. Вони відкриваються назовні парними отворами — дихальцями, або стигмами, а кінцеві гілки обплітають усі внутрішні органи, транспортуючи до них кисень.
Статева система. Багатоніжки — роздільностатеві тварини. Запліднення в них сперматофорне або внутрішнє. Яйця здебільшого багаті на жовток, дробіння поверхневе. Розвиток анаморфний.
Філогенія багатоніжок (за Клюге, 2002)
У підтипі Багатоніжки виділяють 4 класи:
Губоногі (Chilopoda)
Двопарноногі, або Ківсяки (Diplopoda)
Пауроподи (Pauropoda)
Симфіли або Сколопендри (Symphyla)
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ 13. Особливості зовнішньої і внутрішньої будови представників надкласу Шестиногі (Hexapoda)
Питання:
Особливості будови комах.
Типи ротових апаратів.
Розмноження та розвиток комах.
Значення комах.
Клас Комахи (Insecta) включає понад 1 млн видів.
Тіло комах розділене на три відділи: голову, груди та черевце.
Голова утворюється з шести сегментів, що повністю зливаються у дорослої комахи. Перший і третій сегменти не мають придатків, другий несе пару вусиків (антен); на четвертому, п'ятому і шо- стому сегментах розвиваються парні частини ротового апарату — верхні щелепи (мандибули), нижні щелепи (максили) і нижня губа. Вусики за будовою і розміром дуже варіюють і служать важливою систематичною ознакою. Вони можуть мати ниткоподібну, пластинчасту, булавоподібну та інші форми. Розвиток і форма щелеп і нижньої губи залежать від характеру живлення. Виділяють такі основні типи ротового апарату:
гризучий (таргани, жуки, мурашки, личинки метеликів);
гризучо-лижучий (бджоли, джмелі);
колючо-сисний — нижня губа утворює довгий хоботок, а щеле- пи перетворюються на колючі щетини (комарі, тля);
сисний — хоботок розвивається з нижніх щелеп, а верхні повні- стю редукуються (метелики);
лижучий — утворений нижньою губою (мухи).

У багатьох дорослих комах, які не живляться, ротові органи взагалі не розвинені (одноденки).
Комахи здатні рухати головою. Рухи ці в більшості випадків обмежені однією площиною (вертикальною або горизонтальною), і лише богомоли можуть повертати голову в декількох площинах. Груди складаються з трьох сегментів, які несуть три пари членистих кінцівок. Залежно від способу пересування комахи кінцівки за будовою можуть бути (рис.29):
ходильні, або бігальні (паличники, багато жуків);
стрибальні (саранові, коники, блохи);
риючі (капустянки, жуки-карапузи);
хапальні (богомоли);
плавальні (жуки-плавунці).

У багатьох комах до другого і третього сегментів грудей прикріплюються органи польоту — крила. Вони утворюються як вирости покривного епітелію і закріплюються жилками. У більш примітивних видів передні та задні крила однакові за будовою і функцією, яку вони виконують (бабки). У мух і комарів розвивається лише передня пара крил, у віялохвостих — задня. Нерідко передні крила виконують захисну функцію, перетворюючись на надкрилля (клопи, жуки). Багато видів комах повторно втратили крила (блохи, воші), що пов'язано з особливостями їх життєдіяльності. Крила приводяться в рух м'язами, що займають значний обсяг грудного відділу.
Черевце комах утворене різним числом сегментів (не більдше дванадцяти). Воно майже завжди позбавлене кінцівок.
Хітинова кутикула у всіх наземних видів вкрита тоненьким шаром воскоподібної речовини, яка перешкоджає випаровуванню води. На тілі комах є численні волоски, що виконують функцію органів дотику або слуху.
Нервова система побудована за принципом черевного нервового ланцюжка. Головний мозок досягає великого розвитку, особливо у суспільних комах.
Фасеткові очі комах розташовані на першому сегменті голови. Нерідко є ще декілька простих очок (наприклад, у медоносної бджоли). Багатьом видам притаманний гарний зір. Так, бджоли можуть розрізняти форму предметів і деякі кольори (червоний, жовтий, зелений, синьо-фіолетовий, ультрафіолетовий). На відміну від людських, очі бджіл чутливі до поляризованого світла. Усі комахи здатні сприймати коливання температури. Терморецептори локалізовані в них в особливих волосках на члениках ніг. У багатьох комах розвинені органи слуху, утворені волосками наногах і черевці. Дужедобре розвинені органи хімічного чуття і нюху. Смакові рецептори знаходяться на ротових частинах і на лапках. Комахи чутливі до рівня вологості повітря і магнітних полів.
У ротову порожнину впадають протоки слинних залоз. Ротова порожнина веде до стравоходу і далі до шлунка. Залозисті клітини кишечнику виробляють травні ферменти, склад яких визначається характером їжі комахи.
Дихальна система представлена добре розвиненою мережею трубок — трахей. Трахеї відкриваються на поверхню тіла невеликими отворами — дихальцями.
Кровоносна система незамкнена. Безбарвна гемолімфа, циркулююча в порожнині тіла комах, служить для транспортування поживних речовин, продуктів обміну, гормонів. Рух гемолімфи забезпечується скороченнями спинної судини — серця.

Видільна система утворена мальпігієвими судинами — тонкими трубкоподібними виростами кишечнику, їх вільні кінці лежать у порожнині тіла. Розчинні продукти обміну поглинаються клітинами стінок мальпігієвих судин і просвітами проходять у кишку. Пристосуванням для акумуляції води є жирове тіло. Унаслідок окиснення жирової тканини виділяється велика кількість води.
Майже всі комахи роздільностатеві. Чоловічі статеві органи представлені парними сім'яниками, сім'явивідними протоками і копулятивним органом, а жіночі — двома яєчниками і жіночими статевими шляхами. У більшості метеликів і жуків запліднення зовнішньо-внутрішнє. Часто зустрічається Ішутріпшє запліднення. Іноді яйця розвиваються шляхом партеногенезу (тля, паличники).
Практично в усіх комах у життєвому циклі присутня стадія личинки — їм властивий розвиток з перетворенням (рис.30). У разі розвитку з неповним перетворенням личинка веде такий самий спосіб життя, як і доросла особина — імаго, У разі розвитку з повним перетворенням личинка виходить з яйця на ранніх етапах розвитку. На стадії лялечки відбувається руйнування личинкових органів і формування органів імаго.
Ріст комах супроводжується линьками. Доросла комаха не линяє.
Комахи освоїли земну поверхню, ґрунт, повітря. Деякі види повторно заселили прісні водойми (клопи-водоміри, деякі їздці), менше проникли в моря (деякі двокрилі). Розміри комах варіюють у дуже широких межах.
Комахи мають велике значення в медицині як трансмісивні тварини, носії збудників хвороб, а в сільському господарстві як шкідники. Зернових продуктів, які знищуються комахами щорічно в всьому світі, вистачило б для харчування 200 млн. людей.
Для людини комахи являються не тільки ектопаразитами, але й трансмісивними тваринами, наприклад воші, передають сипний та зворотний тиф, а також волинську лихоманку. Блохи переносять – чуму, туляремію, енцефаліт, епідемічний сипний тиф, жовту лихоманку. Двокрилі (мухи, комарі) переносять людині різноманітних протозоонозів, гельмінтозів та інфекцій. Муха це-це переносить трипанозомоз, комар анофелес – малярійних плазмодіїв, домашня муха – яйця аскариди та інших паразитичних червей, а також різноманітних мікробів: поліомієліта, тифа, туберкульоза, холери, сапа, дизентерію та інш. Муха-жигалка – сибірську виразку, туляремію. Личинки багатьох мух являються паразитами людини і тварини, викликають захворення, що називаються міазами тканевими та черевними (личинки мухи Вольфарта).
Корисні комахи відносяться до перетинчастокрилих - оси та бджоли, які є активними запилювачами рослин. Домашня комаха медоносна бджола дає воск та мед. Загальна продуктивність дбжіл Європи складає 10 000 тон
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ. 14. Тип Голкошкі́рі (Echinodermata).
Питання:
1.Анатомія і морфологія.
2.Пристосування до умов життя.
3.Систематика голкошкірих.
ТипГолкошкі́рі (Echinodermata) — другий за кількістю (після хордових) тип вторинноротих тварин, що містить близько 6 300 сучасних видів. Усі вони — виключно морські придонні тварини, що дуже чутливо реагують на опріснення. Майже всі, крім кількох батипелагічних (таких, що мешкають на великих глибинах) голотурій, представляють собою типові бентосні форми. В батіальній та ультраабісальній зонах на частку голкошкірих може приходитися до 90 % загальної біомаси бентосу. Деякі морські зірки та голотурії зустрічаються лише на глибинах більше 2000 м.
Походження.Голкошкірі відомі з раннього Кембрію. Особливо багато вони були представлені в пізньому Палеозої. Тоді ж з'явилася і більшість груп, що дійшли до нас у викопному стані, і є далекими родичами сучасних форм. Завдяки хорошому збереженню їх вапнякових скелетів відомо біля 10 000 викопних видів голкошкірих. З близько 20 таксонів до нашого часу дожило лише шість: морські лілії Crinoida (біля 620 видів), морські зірки Asteroida (близько 1500 видів), офіури або змієхвостки Ophiuroida (близько 2000 видів), морські огірки Holothuroida (приблизно 1200 видів) та морські їжаки Echinoida (біля 950 видів). Шостий таксон Concentricycloida (морські маргаритки), відкритий лише у 1986 році, та відомий по двом видам з глибини біля 1000 метрів у Новій Зеландії та 2000 метрів з Карибського моря, можливо належить до морських зірок.
Голкошкірі, як правило, вільноживучі тварини з радіальною, здебільшого п'ятипроменевою симетрією та елементами білатеральної симетрії. У тілі голкошкірих розрізняють оральну та протилежну аборальну сторони. На оральній стороні розташований ротовий отвір. Деякі викопні форми дають підстави вважати радіальну симетрію вторинним явищем. На це вказує також двобічна симетрія личинок голкошкірих та будова ряду органів сучасних видів.
Ендоскелет.У підшкірному шарі Голкошкірих розвивається біомінеральний ендоскелет, який складається з вапняних платівок і часто утворює різноманітні зовнішні придатки: голки, шипи і педіцеляріі. У багатьох морських їжаків ці придатки розвиваються особливо сильно. Голки виконують захисну функцію. Часто вони бувають рухливими. Деякі морські їжаки направляють голки в бік наближення небезпеки. Це допомагає їм вижити в морях. Педицелляріі — це голки, видозмінені в хапальні щипчики. З їх допомогою тварина чиститься і позбавляється від паразитів. У морських їжаків скелет бере участь у формуванні особливого жувального органу — арістотелевого ліхтаря. У офіура скелет особливо розвинений в променях, де він утворює ряд масивних вапняних члеників — хребців. У голотурій скелет редукований.
Покриви.До складу епідерми Голкошкірих входять механорецепторні клітини, що забезпечують дотик, пігментні клітини, що обумовлюють забарвлення тварини, за допомогою якого можна сховатися від хижака, це допомагає вижити, і залозисті клітини, що виділяють липкий секрет або навіть токсини. Однією з найбільш оригінальних рис будови Голкошкірих слід вважати складну диференціацію целому на ряд систем: амбулакральну і перигемальну системи.
Амбулакральна система унікальна для всього царства тварин. Це сполучена з навколишнім середовищем через кам'янистий канал і мадрепорову платівку мережу каналів, заповнена рідиною, за складом близькою до морської води. Від радіальних амбулакральних каналів відходить безліч амбулакральних ніжок, в основі яких знаходяться ампули — м'язові бульбашки, при скороченні яких ніжка подовжується. На кінці ніжки знаходиться присосок. Амбулакральна система бере участь в диханні, пересуванні та добуванні їжі. Це допомагає їй вижити у морі. Так, за допомогою спільної роботи безлічі амбулакральних ніжок морська зірка може розкрити раковину двостулкової молюска.
Перигемальная система — це сукупність каналів і порожнин (синусів), що оточують кровоносну систему тварини. Кровоносна система слабко розвинена і являє собою систему порожнин в сполучної тканини (лакун), що не мають ендотеліальної вистилки. У кожному промені знаходяться два радіальних перигемальних канали, в перегородці між якими розташовується радіальна кровоносна судина. Радіальні судини впадають у оральне кровоносне кільце, що лежить в перегородці між двома кільцевими перигемальними каналами. Статевий синус оточує аборальне кровоносне кільце і статевої Столон. Два кровоносних кільця пов'язані осьовим органом, оточеним лівим і правим осьовими синусами.
В одному з інтеррадіусів голкошкірих розташований осьовий комплекс органів. До його складу входять органи з різних систем:
кам'янистий канал, який з'єднує кільцевий амбулакральний канал з мадрепоровою платівкою;
осьовий орган, усередині якого розташовується мережа кровоносних судин;
лівий осьовий синус — частина целому, що з'єднує внутрішній кільцевий перигемальний канал з правим осьовим синусом;
правий осьовий синус, здатний ритмічно скорочуватися і цим сприяти руху крові в судинах, тобто виконує функції серця;
статевий синус — ділянка целому, що містить статевий тяж, який складається з незрілих статевих клітин.
Нервова система голкошкірих примітивна, складається з трьох окремих частин, побудованих по радіальному плану: нервове кільце і радіальні нервові тяжі. В оральній стінці тіла лежать дві нервові системи — чутлива і рухова. У аборальній стінці тіла — тільки рухова. Органи чуттів голкошкірих досить різноманітні, але примітивні за будовою. Вони дифузно розподілені по тілу у вигляді різних чутливих клітин (функції дотику, хімічного чуття, зору). Світлочутливі клітини можуть бути зібрані в очах. У морських зірок очі розташовані на кінцях променів, а в морських їжаків — навколо анального отвору.
Розмноження.Більшість голкошкірих — роздільностатеві тварини, утворюють багато дрібних, бідних жовтком яєць і метають їх у воду. Таким чином, запліднення у голкошкірих зовнішнє. Розвиток зародка відбувається у воді, у складі меропланктона. Запліднена яйцеклітина (зигота) починає дробитися і через деякий час утворює бластули. Дроблення повне, радіального типу.
Зазвичай голкошкірі вилуплюються з яйця на стадії бластули. Така бластулоподібна личинка являє собою міхур, стінка якого складається з одного епітелізованого шару джгутиконосних клітин (бластомерів), а порожнина якого (бластоцель) заповнена драглистою рідиною. Бластула здатна пересуватися за допомогою джгутиків. Часто на анімальному полюсі личинки джгутики довші і виконують сенсорну функцію (апікальний орган).
Через деякий час на вегетативному полюсі відбувається виселення (імміграція) деяких клітин в бластоцель. Це клітини первинної мезенхіми, що беруть участь в утворенні личинкового скелета. Після цього відбувається інвагінація вегетативної стінки в бластоцель, в результаті чого формується сліпо замкнута первинна кишка (архентерон). Одночасно з цим з вершини архентерона в бластоцель виселяються клітини вторинної мезенхіми, які володіють амебоїдною рухливістю і сприяють інвагінації, а згодом — відокремлення целому, утворення рота та дорослих рук. Потім відбувається відокремлення целомічної мезодерми, що входить до складу архентерона, шляхом утворення випинання стінки, яке відшнуровується у формі целомічного мішка, тобто ентероцельним способом. Надалі целому ділиться спочатку на правий і лівий, а потім — на три пари целому: праві і ліві аксоцелі (передні), гідроцелі (середні) і соматоцелі (задні). Зазвичай ліві целому випереджають у розвитку праві, що пов'язано з їх провідною роллю в метаморфозиах. На цьому завершується процес гаструляціі, в результаті якого відокремлюються три зародкових листка: ектодерма (личинкові покриви), ентодерми (кишка) і мезодерма (первинна і вторинна Мезенхіма, загалом). Бластопора зміщується на черевну сторону і стає порожниною (анусом). На передньому кінці, на черевній стороні утворюється інвагінація ектодерми, так звана ротова бухта (стомодеум), яке зливається з кишкою і утворює рот. Кишка розчленовується на три відділи: стравохід, шлунок розширений і тонку кишку. Тіло набуває яйцевидної форми. Навколо рота утворюється навколоротова западина, на краю якої формується війковий шнур, на решті поверхні тіла вії зникають. За допомогою биття війок клітин війкового шнура, личинка пересувається і підганяє їжу до рота. Ця спільна для більшості голкошкірих стадія білатерально-симетричною личинки отримала назву діплеврула.
Надалі личинка набуває пристосування до планктонного способу життя, різні у різних класів, і стає плутеусом (морські їжаки і офіури), аурікуляріею (голотурії) або біпінаріею, а пізніше — брахіоляріею (морські зірки). У видів з великою кількістю жовтка з яйця розвиваються лецитотрофні (тобто що харчуються жовтком) бочкоподібні личинки доліоляріі, підперезані поперечними війковими кільцями. Через деякий час вільноплаваючі білатерально-симетричні личинки приступають до метаморфозу, в результаті якого трансформуються в радіально-симетрична доросла тварина. У тілі личинки утворюється зачаток майбутньої дорослої тварини (імагінальний диск). На лівій стороні личинки формується оральна сторона тварини, а на правій — аборальна. До кінця метаморфозу відбувається повна редукція личинкових органів.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,
ЛЕКЦІЯ. 15. Тип хордові (Chordata).
Загальні ознаки хордових.
Характеристика представників класів Ланцетників, Круглоротих.
Значення в еволюції хордових.
Еволюційні особливості будови:
виникнення внутрішнього скелета — хорди, пружного хряща, заміщеного надалі кістковими елементами;
організація нервової системи за типом нервової трубки ектодермального походження;
поєднання дихальної і травної систем: зябра (у водних хордових) і легені (у наземних) утворюються як вирости глотки, а не як похідні кінцівок (у безхребетних).
Усі представники типу — вториннороті тварини. На певному етапі ембріонального розвитку первинний рот (гастропор) заростає ектодермою, а на протилежному кінці зародка утворюється вторинний рот, який і буде ротовим отвором у дорослого організму. У ділянці гастропору знаходитиметься анальний отвір.
Тип Хордові включає три підтипи: Безчерепні; Личинковохордові, або Покривники; Черепні, або Хребетні.
Клас Ланцетники. Клас включає близько ЗО сучасних видів.
Ланцетники — малорухливі тварини, які проводять більшість часу, зарившись у пісок.
Т іло
видовжене, рибоподібне. На спинному
боці вздовж усього
тіла проходить
спинний плавець, який переходить у
хвостовий плавець,
що має форму ланцета.
іло
видовжене, рибоподібне. На спинному
боці вздовж усього
тіла проходить
спинний плавець, який переходить у
хвостовий плавець,
що має форму ланцета.
Шкіра ланцетника складається з двох шарів: зовнішнього — епідермісу, і внутрішнього — дерми. Епідерміс, на відміну від решти хордових, представлений одним шаром клітин, які виділяють тонку кутикулу (рис.31).
Скелет безчерепних утворений хордою — еластичним пружним хрящем. Хорда служить опорою для внутрішніх органів і місцем прикріплення м'язів. Поперечносмугасті м'язи складаються з окремих сегментів.
Нервова система представлена нервовою трубкою, що тягнеться уздовж тіла над хордою. Поділ на мозковий і спинний відділи відсутній. У кожному сегменті від нервової трубки відходять по дві пари нервів.
На передньому кінці тіла знаходиться нюхова ямка, в якій локалізовані нюхові клітини. Уздовж всієї нервової трубки зосереджені органи зору — очки, здатні відрізняти світло від темряви крізь прозорі покриви тіла.
Харчові частинки потрапляють до рота завдяки руху щупалець, що оточують ротову воронку. Рот переходить у глотку, стінки якої прорізані значною кількістю зябрових щілин. Придатнідля живлення часточки обволікаються слизом і надходять у кишечник — коротку пряму трубку. У початковому її відділі є печінковий виріст.
Дихальна система тісно пов'язана з травною. Зяброві щілини, пронизані капілярами, постійно омиваються водою. Щілини відкриваються в атріальну порожнину, яка має один вихідний отвір на нижній поверхні тіла. Вода, потрапляючи в ротовий отвір і далі в глотку, профільтровується крізь зяброві щілини в атріальну порожнину, а звідти крізь вихідний отвір викидається назовні.
Кровоносна система замкнена. Серця немає; його роль виконують пульсуючі кровоносні судини. Цими судинами кров подається до зябер, а далі — до всіх органів тіла. Кров не містить дихальних пігментів.
Система виділення ланцетника подібна до метанефридій кільчастих червів. Одним кінцем звиті трубки відкриваються в целом, а другим — в атріальну порожнину, звідки розчинні продукти виділення видаляються з організму.
Ланцетники роздільностатеві. Статеві залози не мають власних вивідних протоків, гамети виділяються в атріальну порожнину, а звідти — у воду. Запліднення зовнішнє. Унаслідок дроблення заплідненого яйця утворюється личинка. Вона дуже рухлива, активно живиться і служить для розселення. Наявність личинкової стадії — характерна особливість розвитку ланцетника.
Клас круглороті (Cyclostomata). Круглороті мають видовжену вугроподібну форму тіла. Парних плавців немає. Шкірні залози виділяють велику кількість слизу, який оберігає від пошкоджень, полегшує пересування у воді (рис.33).

Скелет представлений хордою і хрящовим черепом.
У круглоротих, на відміну від ланцетників, нервова трубка розділяється на.спинний і головний мозок. У головному мозку виділяють такі відділи:
передній, проміжний, середній, довгастий мозок і мозочок. З органів чуття розвинений нюх, зір (очірозташовані з боків голови під шкірою), орган слуху (внутрішнє вухо), рівноваги (два півкруглі канали) і бічна лінія — орган сприйняття руху й орієнтації в просторі.
Круглороті — напівпаразити. Для присмоктування до жертви служить присмоктувальна воронка, наділена роговими зубчиками. Живляться міноги і міксини кров'ю та м'якими тканинами.
Оскільки круглороті присмоктуються ротовим отвором до жертви, у них виробилися механізми, що дозволяють дихати під час живлення. Зяброві мішки круглоротих знаходяться всередині скелета зябрового апарату — зябрових ґраток. Кожний зябровий мішок має зовнішній і внутрішній отвори (останній відкривається в глотку). Коли тварина вільно плаває, вода надходить у зяброві мішки через рот і внутрішні отвори, а після газообміну виходить через зовнішні отвори. Якщо мінога або міксина присмоктується до риби, вода циркулює тільки через зовнішні отвори.
Кровоносна система замкнена. З'являється серце, яке складається з передсердя і шлуночка. Насичена вуглекислим газом венозна кров надходить у передсердя, далі в шлуночок, а звідти — до зябер. Окиснена артеріальна кров збирається в непарну спинну аорту ,яка лежить під хордою, і розноситься по всьому тілу.
У круглоротих вперше з'являються тулубні (первинні) нир- ки — орган виділення і осморегуляції. Нирка має вигляд стрічки, від якої відходить сечовід, який відкривається назовні сечостатевим отвором.
Круглороті роздільностатеві. Чоловічі статеві залози представлені непарним сім'яником, жіночі — яєчником. Як і в ланцетника, вони не мають власних вивідних протоків. Запліднення зовнішнє. Міноги розвиваються з перетворенням, проходячи личинкову стадію; у міксин розвиток прямий.
Основна література:
1. В.Ф. Натали. Зоология беспозвоночных. М., Главполитграфздат.,
1951.-536 с.,
2. В.А. Догель. Зоология беспозвоночных. М., Высш. школа, 1981.- 606 с.,
3.М.П. Савчук. Зоологія безхребетних. К., Радянська школа, 1965.- 502 с.,
4. А.В. Иванов, Ю.И. Полянский, А.А. Стрелков. Большой практикум по зоологии беспозвоночных. Часть 1. М., Советская наука. 1958.- 558с.
4. П.Б. Гофман-Кадошников, Д.Ф.Петров. Биология с общей генетикой. М.- "Медицина", 1966.-511 с.,
5. Э.Хардон, Р. Венер. Общая зоология. М., "Мир"., 1989.- 523 с.,
6. И.П. Аносов, Л.Я. Кулинич. Основы эволюционной теории. К., “Твір інтер”. 1999.- 286 с.,