

2.5. Модели роста, учитывающие затраты на поддержание жизнедеятельности
Влияние процессов поддержания жизнедеятельности на рост микроорганизмов описывается двумя моделями: «энергии поддержания» и «эндогенного дыхания». Первая из них, предложенная Пертом, получила экспериментальное подтверждение и широко применяется в микробиологии. Вторая была разработана Гербертом для объяснения снижения прироста биомассы в процессе биологической очистки активным илом.
Согласно модели «энергии поддержания» внеклеточный субстрат используется клетками в двух направлениях: на рост и получение энергии поддержания жизнедеятельности:

Соответственно,
общее количество потребленного субстрата
![]() представляется в виде суммы затрат на
рост
представляется в виде суммы затрат на
рост
![]() и поддержание жизнедеятельности
и поддержание жизнедеятельности
![]() :
:
![]() . (2.22)
. (2.22)
Принимается, что затраты субстрата на поддержание жизнедеятельности пропорциональны концентрации биомассы:
![]() , (2.23)
, (2.23)
где
![]() коэффициент энергии
поддержания (удельная скорость потребления
субстрата на поддержание жизнедеятельности).
коэффициент энергии
поддержания (удельная скорость потребления
субстрата на поддержание жизнедеятельности).
Вводится истинный
экономический коэффициент
![]() ,
равный экономическому коэффициенту
при нулевых затратах субстрата на
поддержание жизнедеятельности:
,
равный экономическому коэффициенту
при нулевых затратах субстрата на
поддержание жизнедеятельности:
![]() , (2.24)
, (2.24)
![]() . (2.25)
. (2.25)
При аэробном росте гетеротрофов на простых органических субстратах (сахара, органические кислоты) в оптимальных условиях =0,45 г/гХПК.
Из соотношений (2.22)-(2.25) вытекает зависимость экономического коэффициента от удельной скорости роста:
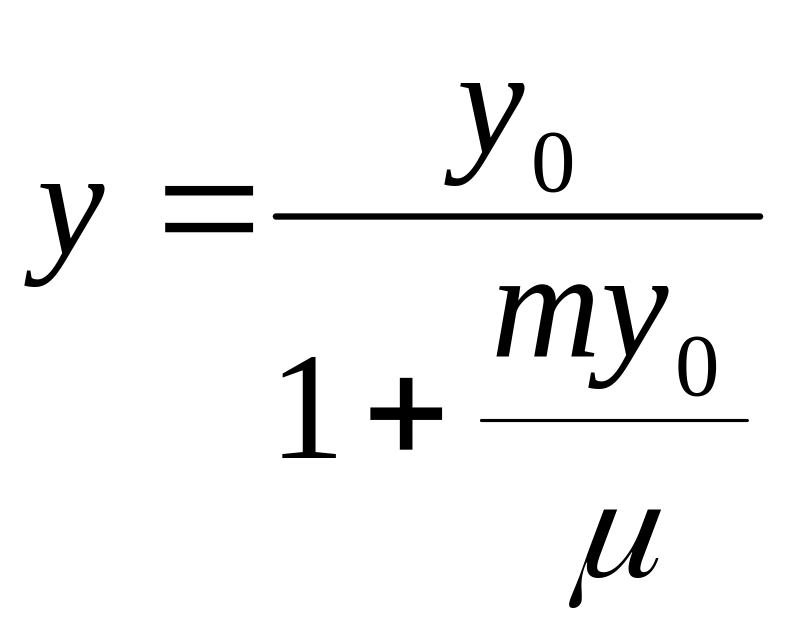 , (2.26)
, (2.26)
![]() . (2.27)
. (2.27)
Потребление кислорода аэробными гетеротрофами описывается аналогичными уравнениями:
![]() , (2.28)
, (2.28)
![]() , (2.29)
, (2.29)
![]() , (2.30)
, (2.30)
где
![]() ,
,
![]() количество кислорода,
потребленного на рост и поддержание
жизнедеятельности;
количество кислорода,
потребленного на рост и поддержание
жизнедеятельности;
![]() истинный энергетический
коэффициент (
=0,4
гО2/гХПК);
истинный энергетический
коэффициент (
=0,4
гО2/гХПК);
![]() удельная скорость
потребления кислорода на процессы
поддержания жизнедеятельности.
удельная скорость
потребления кислорода на процессы
поддержания жизнедеятельности.
Если концентрация органического субстрата измеряется в кислородных единицах (по ХПК), то с учетом (2.6) выполняется:
![]() ;
;
![]() , (2.31)
, (2.31)
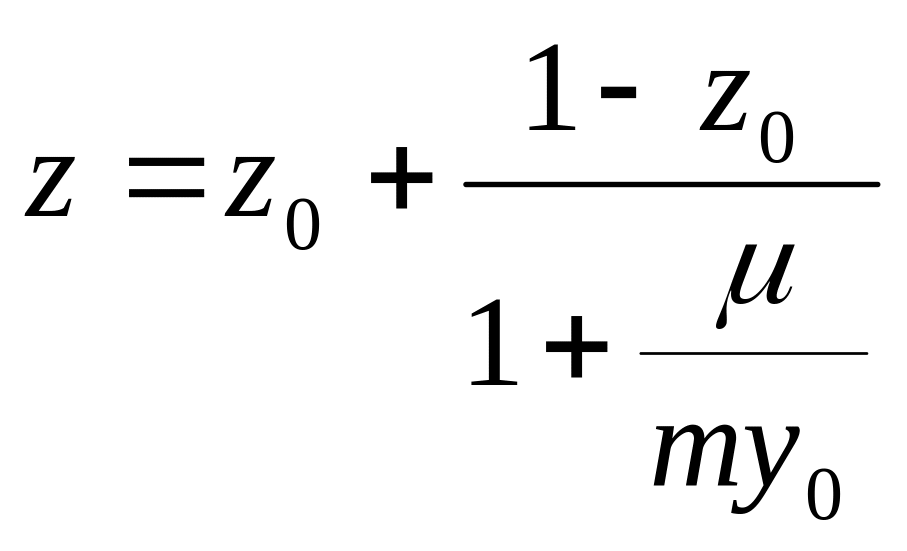 . (2.32)
. (2.32)
Модель эндогенного дыхания постулирует, что энергия поддержания жизнедеятельности образуется в результате самоокисления биомассы по схеме:

Основные уравнения этой модели имеют вид:
![]() , (2.33)
, (2.33)
![]() , (2.34)
, (2.34)
где
![]() константа скорости
самоокисления биомассы;
константа скорости
самоокисления биомассы;
![]() удельная скорость
эндогенного дыхания (удельная скорость
потребления кислорода на самоокисление
биомассы для получения энергии на по
удельная скорость
эндогенного дыхания (удельная скорость
потребления кислорода на самоокисление
биомассы для получения энергии на по
Из (2.33), (2.34) следует:
 , (2.35)
, (2.35)
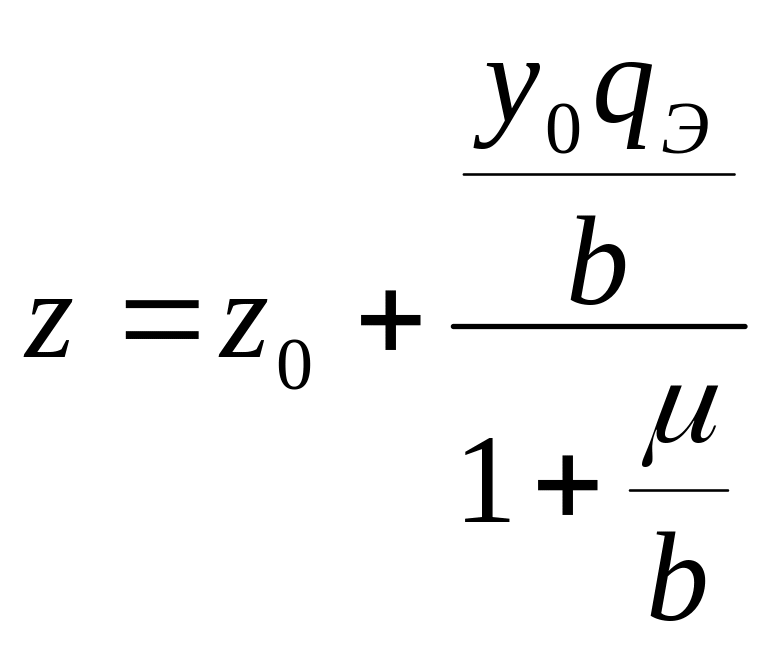 . (2.36)
. (2.36)
Из сопоставления
уравнений (2.26), (2.32) и (2.35), (2.36) видно, что
обе модели дают одинаковые результаты
при
![]() ,
,
![]() ,
где
кислородный эквивалент биомассы.
,
где
кислородный эквивалент биомассы.
Зависимость коэффициентов и от удельной скорости роста показана на рис. 2.6. Для экспериментального определения параметров модели можно использовать линеаризованную форму (2.27).
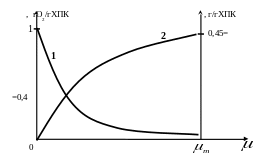
Рис. 2.6. Зависимость энергетического коэффициента (1) и экономического коэффициента (2) от удельной скорости роста
Для бактерий
=0,001-0,1
ч-1.
Поэтому при
![]() ,
когда
,
когда
![]() выполняется:
выполняется:
![]() ,
,
![]() .
Но в условиях лимитирования роста (
.
Но в условиях лимитирования роста (![]() ,
,
![]() )
экономический коэффициент существенно
ниже истинного значения (
)
экономический коэффициент существенно
ниже истинного значения (![]() ),
соответственно
),
соответственно
![]() .
Такая система типична для систем
биологической очистки, задачей которых
является снижение до минимума содержания
органических веществ в воде (
.
Такая система типична для систем
биологической очистки, задачей которых
является снижение до минимума содержания
органических веществ в воде (![]() ).
).
Рассмотренные модели позволяют уточнить уравнение Моно. Из (2.22)-(2.25) и (2.33) в совокупности с уравнением (2.8) следует:
![]() (2.37)
(2.37)
или
![]() . (2.38)
. (2.38)
Уравнения (2.37),
(2.38) отражают тот факт, что рост
микроорганизмов возможен лишь при
концентрации субстрата выше некоторой
минимальной величины
![]() .
При
.
При
![]() весь внеклеточный субстрат расходуется
на поддержание жизнедеятельности и
клетки не растут, а напротив, вынуждены
окислять свои резервы, соответственно
весь внеклеточный субстрат расходуется
на поддержание жизнедеятельности и
клетки не растут, а напротив, вынуждены
окислять свои резервы, соответственно
![]() .
.
Величину легко найти из уравнений (2.37), (2.38) при =0:
![]() . (2.39)
. (2.39)
Так как
![]() (
(![]() ),
то в первом приближении получим:
),
то в первом приближении получим:
![]() . (2.40)
. (2.40)
Отсюда следует:
![]() .
.
Так как для органических субстратов =1-10 мг/л, то составляет всего 0,01-0,1 мг/л. Однако при лимитировании роста по диффузии субстрата величина может быть значительно больше (до нескольких мг/л) и в соответствии с (2.13) определяется по формуле:
![]() , (2.41)
, (2.41)
где задается уравнением (2.14).