

2.2. Затруднения в интерпритации наследования признаков при неполном доминировании.
При дальнейшем изучении передачи потомству родительских признаков было выяснено, что имеется множество вариантов, когда законы Г. Менделя не выполняются. Это отклонения наблюдаются как при внутриаллельном, так и при межаллельном взаимодействии генов.
Затруднения в интерпретации наследования признаков возникают: при плейотропии, наличии летальных генов, неполном доминировании, сверхдоминировании, кодоминировании, множественном аллелизме, аллельном исключении и при сцепленном наследовании.
Плейотропия – это явление, когда один ген отвечает за проявление нескольких признаков. Вероятно, это наблюдается в тех случаях, когда наследуется ген, обеспечивающий недостаточность фермента или структурного компонента, играющих важную роль в формировании нескольких типов тканей или в одной, но широко представленной в организме. Примерами плейотропного действия гена являются синдром Марфана и «голубых склер».
Иногда в плейотропном проявлении гена гомозиготы не жизнеспособны (брахидактилия). Известны случаи, когда летальные гены отвечают и за проявление одного признака (серповидно-клеточная анемия - видоизмененный гемоглобин). Неполное доминирование – это явление, когда доминантный ген не полностью подавляет рецессивный и потомки получают лишь часть доминантного признака. Известно и противоположное явление – сверхдоминирование. В этом случае доминантный ген проявляет себя сильнее в гетерозиготном состоянии, нежели в гомозиготном. Вероятно, это явление можно объяснить взаимодействием продуктов генной активности. Повышение жизнеспособности у потомков с увеличением количества гетерозиготных аллелей получило название гетерозиса.
При кодоминировании гены одной аллельной пары равнозначны, и, находясь в генотипе, оба проявляют свое действие. Типичным примером кодоминирования является наследование групп крови по системе АВО и MNS.
Множественные аллели – это явление, когда в популяции более чем два аллеля. Помимо доминантного и рецессивного генов имеются еще и промежуточные, которые подавляются доминантным геном, а для рецессивного являются доминантными.
Аллельное исключение наблюдается у гетерозиготных организмов, когда в одних клетках активна одна аллель, а в других – другая. Это внутриаллельное взаимодействие присутствует у всех женщин, т.к. в одних клетках спирализуется отцовская Х хромосома, а в других – материнская.
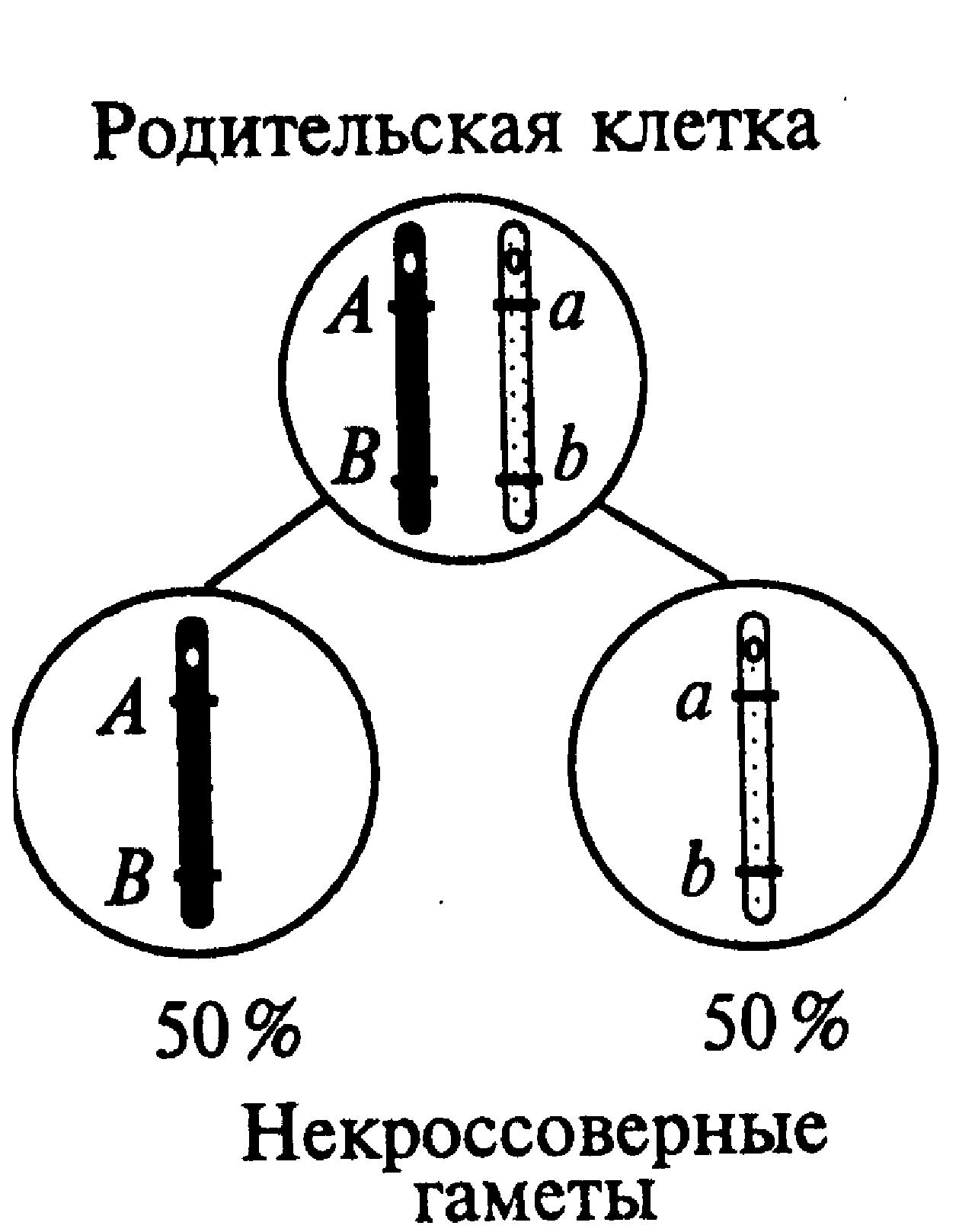
Сцепленное наследование генов. Особенности совместного наследования генов одной пары гомологичных хромосом (сцепленных генов) были изучены Т. Морганом и его сотрудниками с помощью дигибридных и тригибридных скрещиваний дрозофилы. Такие гены преимущественно наследуются сцепленно, однако эта тенденция с той или иной вероятностью может нарушаться за счет кроссинговеров, приводящих к генетическому обмену на разных участках гомологичных хромосом. Возможны два типа сцепления генов — полное и неполное (частичное). Полное сцепление было обнаружено в лаборатории Т. Моргана у самцов дрозофил. Выяснили, что в этом случае отсутствовал кроссинговер при конъюгации гомологичных хромосом в профазе первого деления мейоза. Так, у дигетерозиготной особи с условным генотипом АВ//аb происходило образование двух вариантов некроссинговерных гамет (АВ и аb) с равной вероятностью (1/2, или 50 %). Полное сцепление должно наблюдаться также и в отношении неаллельных генов негомологичных участков половых хромосом у организмов гетерогаметного пола (рис.14).
|
|
Рис. 14. Образование гамет при полном сцеплении генов |
Рис. 15. Образование гамет при неполном сцеплении генов |
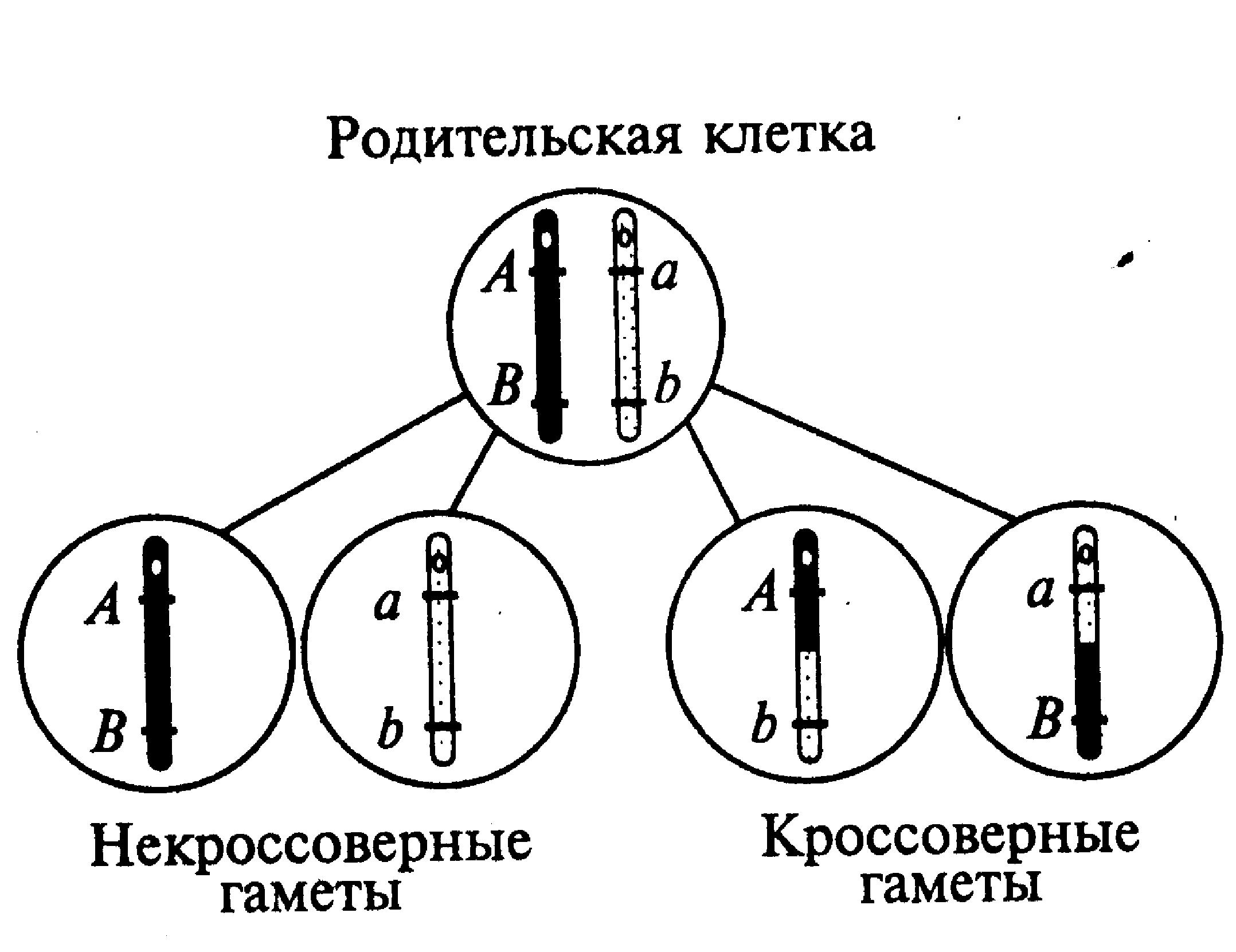
Гораздо чаще встречается неполное (частичное) сцепление генов, в основе которого лежит генетическая рекомбинация (кроссинговеры) на тех или иных участках гомологичных хромосом во время мейотического деления клеток. Как видно на рис.15 у индивидуума с тем же условным генотипом АВ//аb формируются четыре варианта гамет — некроссинговерные (АВ, аb) и кроссинговерные гаметы (Аb, аВ). Поскольку кроссинговер представляет собой случайное и относительно редкое событие, то количество образовавшихся с его участием кроссинговерных гамет будет меньшим, чем количество некроссинговерных гамет. В сумме вероятность последних в каждом отдельном случае всегда больше, чем 0,5 (50%).
В соответствии с правилом, установленным Морганом, количество кроссинговеров на участках между локусами сцепленных генов пропорционально расстояниям между этими локусами на хромосомах. Следовательно, чем больше расстояние между двумя конкретными локусами, тем более значительным будет число различных кроссинговеров на участках двух гомологичных хромосом, расположенных между этими локусами, и более значительным количество соответствующих кроссинговерных гамет. Нужно, однако, иметь в виду, что на эффективность самого процесса рекомбинации у разных организмов могут оказывать влияние как их генетические особенности, так и некоторые факторы среды. Поэтому возможны, например, те или иные различия в частоте кроссинговеров на одном и том же хромосомном участке для ивидуумов, относящихся к одному виду, но имеющих разную половую принадлежность. При этом более низкая частота обычно характерна для особей гетерогаметного пола, а у самцов дрозофил в половых хромосомах кроссинговеры вообще не происходят.
Для количественной оценки кроссинговеров между локусами конкретных генов и соответствующих кроссинговерных гамет проводят анализирующие скрещивания особей с определенными различиями по двум либо трем парам сцепленных аллелей. В таких скрещиваниях анализу подвергается генетическая структура одного из родителей, имеющего доминантные гены (признаки), интересующие исследователя, тогда как второй родитель является гомозиготным по рецессивным аллелям изучаемых генов. В качестве примера рассмотрим дигибридное анализирующее скрещивание дрозофил, проведенное Т.Морганом и его сотрудниками с использованием дигетерозиготной самки, в генотипе которой имелись доминантные гены серой окраски тела (условный символ А) и длинных (нормальных) крыльев (символ В), и дигомозиготного самца с рецессивными аллелями черной окраски (а) и коротких («зачаточных») крыльев (b). Это скрещивание можно представить в виде схемы:
Р |
|
G |
АВ, аb, Аb, аВ аb |
F1 |
АВ //аb: аb//аb: Аb//аb : аВ//аb |
При исследовании полученного потомства были обнаружены четыре фенотипических варианта со следующими частотами: АВ (серый цвет, длинные крылья) — 41,5%, аb (черный цвет, короткие крылья) — 41,5%, Аb (серый цвет, короткие крылья) — 8,5 % и аВ (черный цвет, длинные крылья) — 8,5%. Таким образом, суммарная частота кроссинговерных фенотипов в потомстве (Ab + аВ) в этом скрещивании составила 17 %. Следовательно, количество соответствующих кроссинговерных генотипов потомства и кроссинговерных гамет самки также можно оценить величиной в 17%. По предложению Т. Моргана при составлении генетических карт дрозофилы расстояния между локусами сцепленных генов стали обозначать в виде количества процентов рекомбинационных событий на участках хромосом между этими локусами. Так, в рассмотренном примере расстояние между локусами генов окраски тела и длины крыльев дрозофилы на хромосомной карте будет определяться величиной в 17% рекомбинации. В дальнейшем 1% рекомбинации стали называть морганидой (сантиморганидой).
Исследование явления сцепления генов позволило Моргану сформулировать главную генетическую теорию — хромосомную теорию наследственности. Ее основные положения выглядят следующим образом: