

С.П. Сапожников Е.М. Лузикова О.И. Московская
Общая и медицинская генетика
Учебное пособие
Чебоксары 2008
Министерство образования и науки
Российской Федерации
ФЕДЕРАЛЬНОЕ АГЕНТСТВО
Федеральное государственное образовательное учреждение высшего профессионального образования
«Чувашский государственный университет
им. И.Н. Ульянова»
С.П. Сапожников Е.М. Лузикова О.И. Московская
Общая и медицинская генетика
Учебное пособие
Чебоксары 2008
Издательство Чувашского университета
УДК
С 195
Рецензенты:
проректор по науке ГОУ ВПО «Чувашский государственный педагогический университет им. И.Я. Яковлева», д-р вет. наук, профессор А.А. Шуканов
канд. биол. наук, доцент кафедры медицинской биологии медицинского института ФГОУ ВПО «Чувашский государственный университет им. И.Н. Ульянова» И.Г. Зеленова
Сапожников С.П.
С 195 Общая и медицинская генетика: учеб. пособие / С.П. Сапожников, Е.М. Лузикова, О.И. Московская.-Чебоксары: Изд-во Чуваш. ун-та, 2008.-195 с.
Учебное пособие включает семь теоретических разделов: «Вопросы молекулярной генетики», «Закономерности наследования признаков. Внутриаллельные отклонения от законов наследования», «Взаимодействие неаллельных генов», «Биология и генетика пола», «Методы исследования в генетике человека. Кариотип», «Изменчивость, ее виды и механизмы», «Популяционная генетика». В учебном пособии собраны новейшие достижения генетики, приводятся практические примеры по тематикам, даны задания для освоения теоретического материала и примеры их решения.
Пособие предназначено для студентов медицинских вузов и биологических факультетов.
Утверждено Редакционно-издательским советом университета в качестве учебного пособия
Отв. редактор: д-р биол. наук, профессор В.Е.Сергеева
ISBN © С.П. Сапожников, Е.М. Лузикова, О.И. Московская, 2008 УДК
Введение
Место генетики среди биологических дисциплин и особый интерес к ней определяется тем, что она изучает основные свойства живых организмов – наследственность и изменчивость, а также способы управления ими. Генетика как наука одной из первых вошла в разряд точных биологических специальностей благодаря открытию дискретных единиц наследственности – генов и методу генетического, в первую очередь гибридологического, анализа. В логике генетического анализа присутствует формализованный подход, позволяющий выдвигать рабочие гипотезы и проводить анализ эмпирических данных на основе изучения расщеплений по признакам в поколениях, взаимодействия аллелей и генов, а также позволяющий выявлять влияние мутагенных и модифицирующих факторов на характер проявления признаков.
Генетика является теоретической основой селекции новых продуктивных форм микроорганизмов, синтезирующих антибиотики, витамины и другие биологически активные соединения, используемые в медицинской промышленности и биотехнологии, а также сельскохозяйственных растений и животных. Область генетических знаний, имеющих отношение к медицине, получила название «медицинская генетика». Она интенсивно расширяется в разных направлениях: изучение генома человека, цитогенетика, молекулярная и биохимическая генетика, иммуногенетика, генетика развития, популяционная генетика, клиническая генетика.
Образование по медицинской генетике включает основы общей генетики (менделизм, учение о хромосомах, химические основы наследственности), основные положения генетики человека и клиническую генетику.
В последнее время происходит быстрое развитие генетики человека, изучающей основные закономерности его наследственности и изменчивости, включая особенности геномной организации, структурно-функциональную организацию отдельных генов, механизмы их наследования и др. В рамках завершающихся исследований по международной программе «Геном человека» совместными усилиями ученых из разных стран составлены генетические карты хромосом человека, содержащие в сумме около 32 000 генных локусов.
Особенности современного этапа генетического изучения человека определяются широким использованием молекулярно-генетических, биохимических, цитогенетических и других новейших методов исследования. Это привело к значительному прогрессу в знаниях о наследственности и изменчивости человека, в том числе о молекулярных механизмах генетических процессов, лежащих в основе нормальных и патологичеких состояний организма.
Знание генетики необходимо врачу любой специальности и биологам всех профилей для понимания механизмов индивидуального развития и его нарушений, природы любого заболевания, рационального подхода к диагностике, лечению и профилактике болезней.
Раздел I. Теоретические основы общей и медицинской генетики. Глава 1. Вопросы молекулярной генетики
1.1. Структура и функции нуклеиновых кислот. Строение молекул днк и рнк
Молекула нуклеиновой кислоты представляет собой полимер (полинуклеотид), состоящий из последовательно соединенных друг с другом мономеров (нуклеотидов). В свою очередь, каждый нуклеотид представляет собой соединение, в котором присутствуют различные молекулы: остаток фосфорной кислоты (фосфат), углевод (пентоза) и азотистое основание (пуриновое либо пиримидиновое).
Н
Остаток
фосфорной кислоты
Остаток
пентозы (рибоза или дезоксирибоза)
Азотистое
основание
(пурин
или пиримидин)
Рис.1. Варианты
изображения строения нуклеотида:
1'-5'— атомы углерода
в молекуле углеводорода (пентозы)
В
Рис. 2. Строение
нуклеозида
(сахарофосфатный
остов)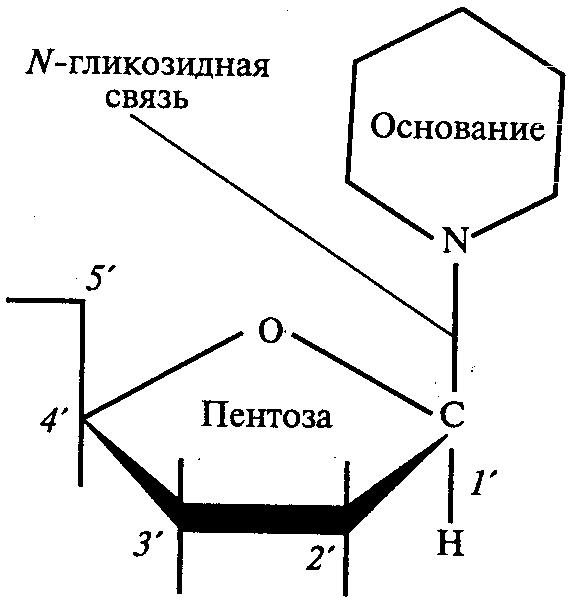
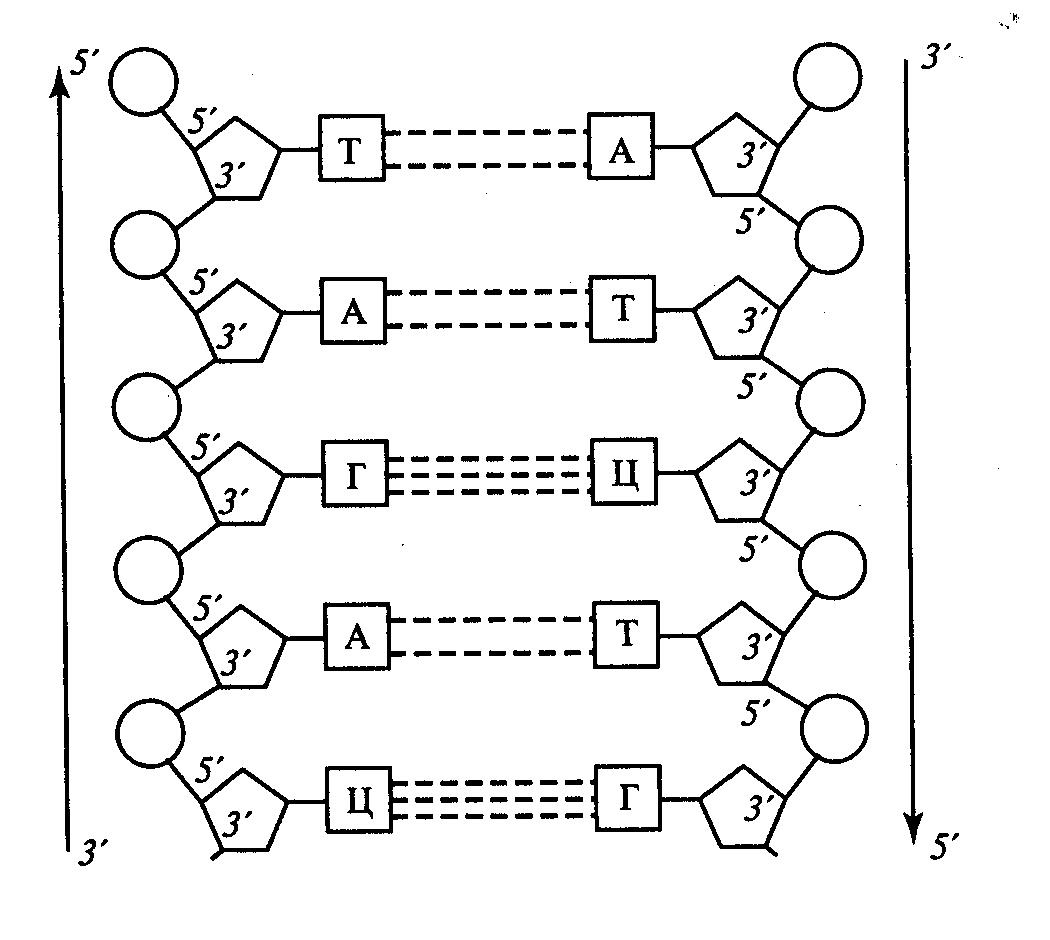
Согласно модели Уотсона и Крика молекула ДНК состоит из двух полинуклеотидных цепочек, соединенных друг с другом с помощью поперечных водородных связей между азотистыми основаниями по комплементарному принципу. Так, аденин одной цепочки соединяется двумя водородными связями с тимином противоположной цепочки, а гуанин и цитозин разных цепочек соединены друг с другом тремя водородными связями. При этом две полинуклеотидные цепочки одной молекулы являются антипараллельными, т.е. напротив 3'-конца одной цепочки находится 5'-конец другой цепочки и наоборот.
На основании данных рентгеноструктурного анализа ДНК Дж. Уотсон и Ф. Крик сделали заключение, что двухцепочечная молекула имеет форму спирали, закрученную слева направо (А – форма, или вторичная структура) (рис. 3).
|
|
Рис. 3. Схема структуры участка молекул ДНК |
1.2. Репликация ДНК.
Расшифровка структуры молекулы ДНК помогла объяснить и принцип ее репликации (удвоения) в клетке. Этот принцип состоит в том, что каждая из двух полинуклеотидных нитей молекулы ДНК служит в качестве матрицы для синтеза новой (комплементарной) нити. Такой принцип репликации ДНК был назван полуконсервативным (рис. 4). В соответствии с этим принципом нуклеотидная последовательность матричной (родительской) нити считывается в направлении 3'→ 5', а синтез новой (дочерней) нити идет в направлении 5'→ 3 '. Поскольку две комплементарные цепочки родительской молекулы ДНК являются антипараллельными, то синтез новой полинуклеотидной цепочки на каждой из них идет в противоположном направлении форме.
У
Рис. 4. Принцип
полуконсервативной репликации ДНК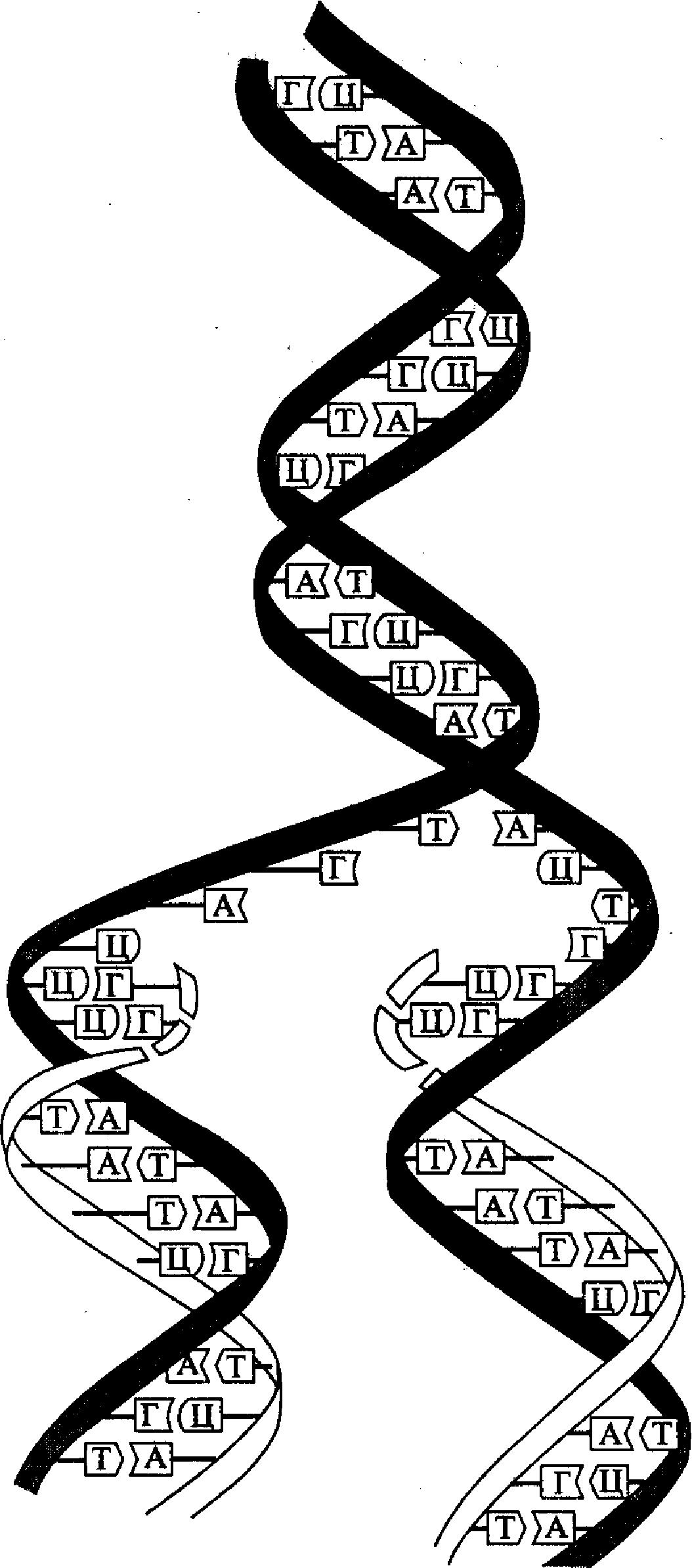
В механизме репликации ДНК у эукариот можно условно выделить три последовательных этапа, которые происходят в репликоне.
I этап связан с быстрым раскручиванием двух полинуклеотидных нитей спирализованной молекулы ДНК на определенном ее участке (в границах работающего репликона) и с их разделением путем разрушения водородных связей между парами комплементарных оснований. При этом образуются два одноцепочечных фрагмента родительской молекулы, каждый из которых выступает в роли матрицы для синтеза комплементарной (дочерней) нити.
Этот этап инициируется в соответствующей точке начала репликации и обеспечивается комплексным участием нескольких различных белков. В результате их действия формируется Г-образная структура, названная вилкой репликации, в которой две родительские цепочки ДНК уже отделены друг от друга. Образовавшаяся вилка репликации быстро продвигается вдоль двойной спирали родительской молекулы ДНК благодаря активности «расплетающего» фермента ДНК - геликазы и при участии группы дестабилизирующих белков (рис. 5). Дестабилизирующие белки связываются только с одноцепочечными (уже раскрученными и разделенными) участками молекулы, препятствуя возникновению на них вторичных складчатых образований («шпилек») за счет случайных соединений между комплементарными нуклеотидами однонитевой структуры. Этот процесс способствует выпрямлению однонитевых участков молекулы, что необходимо для нормального выполнения ими матричных функций.
Быстрое расплетание ДНК с помощью геликазы без дополнительного вращения нитей по отношению друг к другу должно приводить к образованию новых витков (узлов) на участках родительской молекулы перед движущейся вилкой репликации, создающих повышенное топологическое напряжение на этих участках. Такое напряжение устраняется еще одним белком ДНК —-топоизомеразой, который, перемещаясь вдоль двухспиральной родительской ДНК перед вилкой репликации, вызывает временные разрывы в одной из цепочек молекулы, разрушая фосфодиэфирные связи и присоединяясь к разорванному концу. Возникший разрыв обеспечивает последующее вращение нити двойной спирали, что в свою очередь приводит к расплетанию образующихся супервитков (узлов). Разрывы полинуклеотидной цепочки, вызванные топоизомеразой, носят обратимый характер, а разорванные концы после раскручивания быстро воссоединяются после разрушения комплекса этого белка с разорванным концом.
Н
Рис. 5. Схема
образования
репликационной
вилки ДНК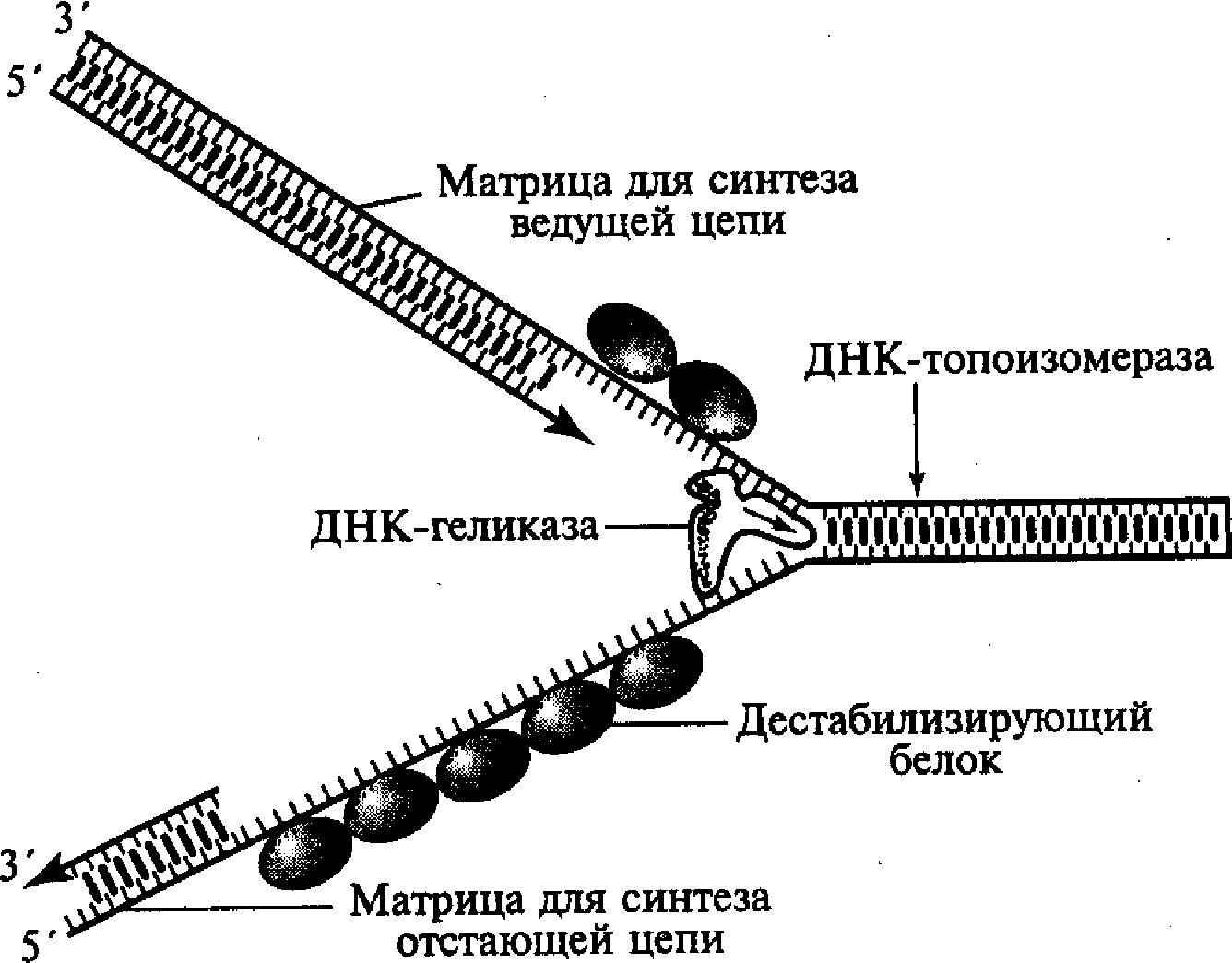
С учетом данного обстоятельства становится понятным асимметричный характер функционирования вилки репликации. Как видно из приведенных схем, на одной из матричных нитей вилки (3'- 5') идет быстрый и непрерывный синтез дочерней нити в направлении 5'→3'. В это же время на другой матрице (5'→3') идет более медленный и прерывистый синтез цепочки с образованием коротких фрагментов по 100-200 нуклеотидов. Эти вновь синтезируемые фрагменты получили название фрагментов Оказаки. Как видно, их синтез идет также в направлении 5'→3'. Считается, что синтез ведущей и отстающей цепочек осуществляют ДНК-полимеразы разных типов. Свободный 3'-конец, необходимый для начала синтеза фрагмента Оказаки, обеспечивается короткой нитью РНК, состоящей примерно из 10 нуклеотидов и получившей название РНК-праймера или РНК-затравки, которая синтезируется с помощью фермента РНК-праймазы (рис. 6). РНК-праймеры могут комплементарно спариваться сразу с несколькими участками на матричной нити ДНК, создавая условия для одновременного синтеза нескольких фрагментов Оказаки при участии ДНК-полимеразы III. Когда синтезированный фрагмент Оказаки достигает 5'-конца очередного РНК-праймера, начинает проявляться 5'-экзонуклеазная активность ДНК-полимеразы 1, которая последовательно выщепляет нуклеотиды РНК в направлении 5'-3'. При этом удаляемый РНК-праймер замещается соответствующим фрагментом ДНК.
I
Рис.6. Синтез
ведущей и отстающей цепей ДНК в области
репликационной вилки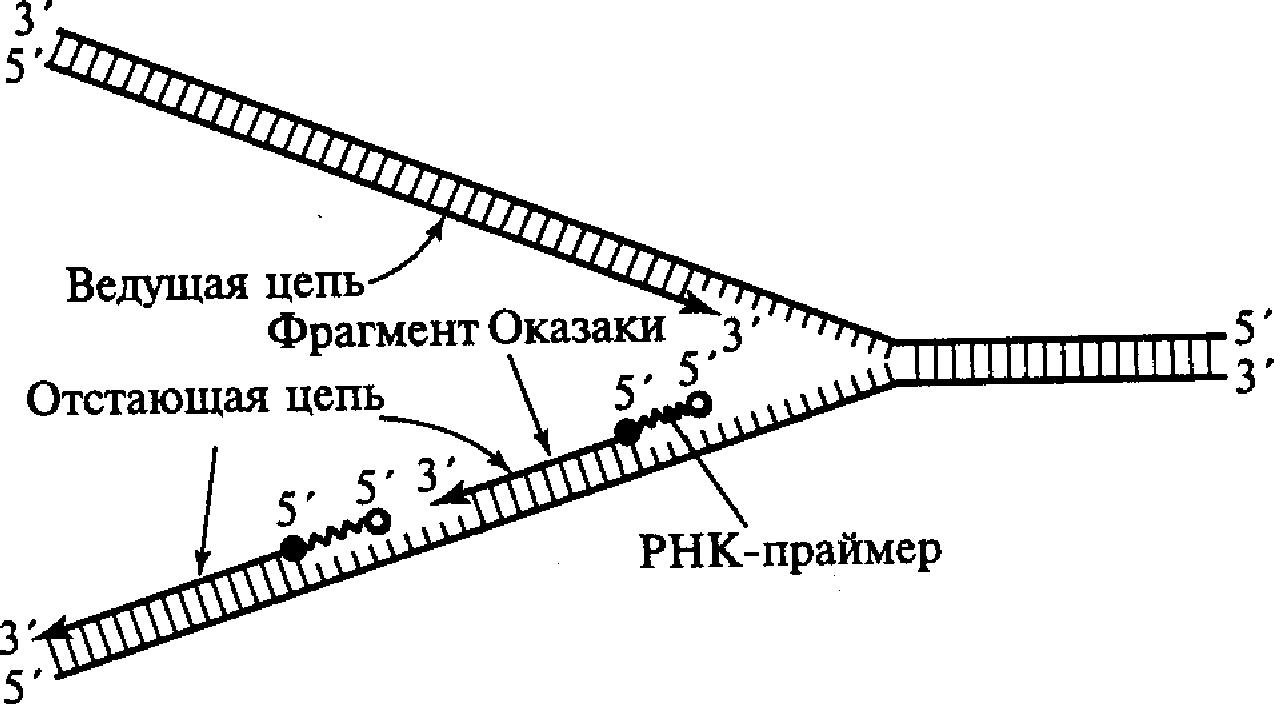
Полирепликонный принцип организации молекулы ДНК различных эукариот, в том числе человека, обеспечивает возможность последовательного копирования генетического материала без одновременного раскручивания всей огромной по размерам и сложно упакованной молекулы, что значительно сокращает время ее репликации. Иными словами, в тот или иной момент времени в одной группе репликонов молекулы процесс копирования может быть уже завершен объединением и спирализацией соответствующих участков, тогда как в другой группе он только начинается расплетанием двухнитевых структур.