

3 Наумов н. П. Я др.1 ч. 3
65

Рис. 29. Загальна схема терморегуляції в гомойотермных організмів (по Слоннму)
окислювання накопичується в аденозинтрифосфорной кислоті, цо забезпечує роботу м'язів; при її розпаді виділяється тепло. Однак при сильному охолодженні такий фосфориліаційний тип окислювання не забезпечує виділення достатньої кількості тепла. Тоді включається вільний або прямий тип тихорєцького подиху (без участі аденозинфосфорних кислот), при якому більша частина вивільнюваної енергії виділяється у вигляді тепла. При виснаженні енергетичних запасів (жири, вуглеводи) і неможливості їхнього поповнення теплопродукція падає й теплокровний організм гине; при цьому смерть наступає не від переохолодження, як звичайно думають, а від виснаження.
При зменшенні маси тіла його відносна поверхня зростає, збільшуючи тепловтрати. Тому при подібних умовах дрібні птахи й ссавці повинні витрачати на підтримку температури тіла більше енергії, чим великі види (мал. 30). Це «правило поверхні» у загальній формі застосовується до всіх гомойотермних тварин. Тому дрібні види споживають відносно більше їжі й кисню, чим більші види. Однак варто враховувати, що на інтенсивності теплопродукції, необхідної для підтримки температури тіла на певному рівні, позначаються, крім розмірів, багато інших морфоекологічних особливостей даного виду: форма тіла, стан теплоізоляційних покривів, рухливість, харчова спеціалізація й доступність кормів, їхня калорійність, добова й сезонна ритміка активності, характер і мікроклімат місць перебування і т.д.
к У
регуляції тепловтрат велику роль грає
фізична терморегуляція. Змінюючи
положення волосяного (пір'яного) покриву,
тварина збільшує або зменшує товщину
повітряного шару й відповідно втрати
тепла. Підвищення теплоізоляційних
властивостей покривів узимку забезпечує
осіннє линяння, під час якого одягається
більш довгий і густий волосяний (пір'яний)
покрив. У песців він настільки збільшує
теплоізоляцію, що дозволяє навіть узимку
не підвищувати споживання кисню, тобто
задовольнятися колишньою кількістю
їжі. Зміна просвіту периферичних
кровоносних судин і швидкості кровотока
також змінює тепловіддачу. Втрати тепла
знижуються при зростанні відкладень
жиру в сполучнотканному шарі шкіри
(особливо характерно для водних тварин).
Частіше дихання й збільшення випару з
поверхні дихальних шляхів сприяє віддачі
надлишку тепла й запобігає перегріву;
у ссавців цьому ж служать потові залози.
Значну роль у терморегуляції грає
поведінка: використання різноманітних
притулків, ночівлі під снігом і багато
чого іншого. Характерні для ряду птахів
і ссавців сезонні кочівлі й міграції
дозволяють їм протягом усього року бути
в районах, сприятливих кліматичних умов
і запасом корму.
У
регуляції тепловтрат велику роль грає
фізична терморегуляція. Змінюючи
положення волосяного (пір'яного) покриву,
тварина збільшує або зменшує товщину
повітряного шару й відповідно втрати
тепла. Підвищення теплоізоляційних
властивостей покривів узимку забезпечує
осіннє линяння, під час якого одягається
більш довгий і густий волосяний (пір'яний)
покрив. У песців він настільки збільшує
теплоізоляцію, що дозволяє навіть узимку
не підвищувати споживання кисню, тобто
задовольнятися колишньою кількістю
їжі. Зміна просвіту периферичних
кровоносних судин і швидкості кровотока
також змінює тепловіддачу. Втрати тепла
знижуються при зростанні відкладень
жиру в сполучнотканному шарі шкіри
(особливо характерно для водних тварин).
Частіше дихання й збільшення випару з
поверхні дихальних шляхів сприяє віддачі
надлишку тепла й запобігає перегріву;
у ссавців цьому ж служать потові залози.
Значну роль у терморегуляції грає
поведінка: використання різноманітних
притулків, ночівлі під снігом і багато
чого іншого. Характерні для ряду птахів
і ссавців сезонні кочівлі й міграції
дозволяють їм протягом усього року бути
в районах, сприятливих кліматичних умов
і запасом корму.
Терморегуляція птахів і ссавців формується в ході індивідуального розвитку (онтогенезу). У частини видів ефективна хімічна терморегуляція встановлюється вже в перші дні після народження (вилуплення), в інших - значно пізніше, і дитинчата на якийсь час виявляються пойкілотермними. Будівля гнізд, нір й інших притулків й обігрів батьками створюють сприятливий для росту й розвитку мікроклімат.
Теплокровність забезпечується високим рівнем обміну речовин: при низьких температурах метаболізм птахів і ссавців у десятки разів вище, ніж у пойкілотермних хребетних. Стійкість «внутрішнього клімату» - необхідна умова безперебійної роботи всіх систем організму, у тому числі центральної нервової системи й органів почуттів. У свою чергу це збагатило сприйняття зовнішнього середовища й забезпечило більш тонке й гнучке пристосування до нього зміною поведінки. Посилення й ускладнення поведінки й розмаїтість зв'язків між особинами вдосконалювало популяційну структуру, дозволяло швидше знаходити їжу, уникати хижаків, а будівництво гнізд, нір, троп, гребель давало можливість активно пристосовувати середовище до своїх потреб.
Теплокровність (гомойотермія), складна вища нервова діяльність, різноманітні форми турботи про потомство, розмаїтість кормів й інші особливості дозволили птахам і ссавцям заселити практично всю Земну куля, включаючи безводні пустелі й найсуворіші гірські райони, утворювати там стійкі поселення й зайняти пануюче положення не тільки у всіх біоценозах суши, але й у частині морів. Це обумовило значення обох класів у житті біосфери й у глобальному круговороті речовин.
Треба ще раз підкреслити, що еволюція птахів і ссавців протікала паралельно, на різній морфофізіологічній основі. Однак загальний напрямок еволюційних перетворень і характер досягнутих біологічних результатів у значній мірі виявився подібним.
КЛАС ПТАХи - AVES
Характеристика класу
Птахи — це покриті пір'ям гомойотермні амніоти, передні кінцівки яких перетворилися в крила. За багатьма морфологічними ознаками вони подібні із плазунами. Ця подібність виражена в наявності рогових лусок хоча б на пальцях задніх кінцівок й у роговому покриві дзьоба, у майже повній відсутності шкірних залоз. Череп діапсидного типу, але зі скороченою верхньою дугою; є лише один потиличний мищелок і подібний із плазунами набір костей. У кінцівках утворюються інтеркарпальні й інтертарзальні зчленування. Ребра несуть крючковидні відростки. Артеріальна кров із серця йде по правій дузі аорти, а схема периферичної кровоносної мережі близька до рептилій. Подібна будова сечостатевої системи й характер ембріонального розвитку.
Однак завдяки багатьом, але найчастіше невеликим морфологічним перетворенням, що торкнулися практично всіх системи органів й забезпечивши значну інтенсифікацію фізіологічних процесів, був істотно підвищений загальний рівень життєдіяльності й створена можливість польоту при збереженні здатності пересуватися й по суші, і в кронах дерев, а в частині видів - у воді й по її поверхні. Стало значно складніше й різноманітніше поведінка, внутрішньовидові відносини й зв'язки з навколишнім середовищем.
Специфічні риси організації птахів, що відрізняють їх від плазунів, різноманітні. Тіло покрите пір'ям, що виконує теплоізоляційну функцію й забезпечує обтичність тіла; пір'я утворюють несучі площини в польоті (крила, хвіст). Перетворення передніх кінцівок у крила супроводжувалося перебудовою кістяка й мускулатури кінцівок і плечового пояса. Одночасні перетворення кістяка й мускулатури задніх кінцівок і тазового пояса забезпечили можливість двоногого ходіння по твердому субстраті й плавання. Пневматизація кісток збільшила їхню міцність. Гетероцільний тип хребців різко збільшив рухливість шиї й голови. Інтенсифікація зору й слуху, прогресивний розвиток головного мозку розширили можливості орієнтування в просторі, поліпшили координацію складних рухів, забезпечили ускладнення поведінки, у тому числі й різкому зростанні ролі індивідуального досвіду, що приводить до вироблення складних умовних рефлексів. Формування здатного до складних рухів дзьоба, незважаючи на редукцію зубів, сприяло різноманітній харчовій спеціалізації й розширило спектр використовуваних кормів, а перебудови в травній трубці (відокремлення мускульного шлунка, подовження кишечнику) поліпшили засвоєння їжі. Утворення пов'язане з легенями системи повітряних мішків інтенсифікувало подих. Повний поділ великого й малого кіл кровообігу сприяло кращому постачанню тканин киснем і поживними речовинами й видаленню вуглекислоти й продуктів розпаду.
Інтенсифікація подиху, кровообігу й травлення забезпечили зростання рівня обміну речовин, що разом з появою теплоізоляційного покриву привело до становлення гомойотермії: підвищилася стійкість птахів до коливань температури зовнішнього середовища й розширилися їхні можливості до розселення. Висиджування яєць і вигодовування пташенят батьками скоротило тривалість ранніх етапів онтогенезу й знизило ембріональну й постембріональну смертність.
Гомойотермія, висока рухливість, складна й різноманітна поведінка - все це дозволило птахам широко розселитися по земній кулі й заселити практично всі наземні місцеперебування. Частина видів успішно освоїла й наземно-водний спосіб життя. Однак глибока спеціалізація до польоту привела до обмеження адаптивної радіації класу птахи. Серед них немає видів з підземним і чисто водним способом життя. Відносно невеликі розходження в розмірах. Відсутність такої вузької спеціалізації дозволило ссавцям освоїти всі типи місцеперебувань (вони не завжди можуть заселяти дрібні океанічні острови), і крім цього заселяти ґрунт (підземні види) і воду; чисто водні форми (китоподібні, сирени) повністю втрачають зв'язки із сушею.
Походження й еволюція птахів
Палеонтологічні матеріали по птахам дуже вбогі. Безсумнівно, що птахи відокремилися від архозаврів – Агсhosauria – що господарювали у мезозойську еру численної й різноманітної групи діапсидних плазунів. Безпосередніх предків птахів потрібно шукати не серед літаючих ящерів, а серед найбільш древньої групи архозаврів - псевдозухій, або текодонтів (Рseudosuchia, seu Thecodontia), які дали початок й іншим групам архозаврів. Псевдозухій по зовнішньому вигляду нагадували ящірок і вели наземний спосіб життя. У частини видів задні кінцівки були трохи довше передніх, і при швидкому бігу тварини, імовірно, зпиралися об землю тільки задніми кінцівками. У деяких особливостях кістяка псевдозухій можна роздивитися риси подібності із птахами (деталі будови черепа, таза, задніх кінцівок).
Проміжні форми між псевдозухіями й птахами, які показували б послідовні стадії розвитку пірйового покриву й перетворення кістяка, поки не виявлені. Припускають, що деякі псевдозухії поступово переходили до деревного способу життя. Розростання рогової луски з боків тіла й хвоста, по задньому краї кінцівок дозволяло подовжити стрибки з гілки на гілку (гіпотетична стадія проптаха, мал. 32). Подальша спеціалізація й добір привели до формування скелетно-м'язової структури й пір'я крила (і відповідних перетворень в інших системах органів), що забезпечило можливість спочатку планіруючого, а потім й активного польоту. Пір'я тіла, можливо, розвивалося спочатку як пристосування до термоізоляції, а вже потім придбало й додаткову функцію забезпечення обтичності тіла; воно сформувалося -

Рис. 32. Гіпотетичний предок птахів - проптах (по Хейльману, 1926)
ще до придбання здатності до польоту. У деяких псевдозухій на тілі були подовжені луски з поздовжнім гребенем і дрібними поперечними ребрами. Мабуть, з них шляхом розчленовування й утворилося перо.
Відокремлення птахів від плазунів, імовірно, відбулося вже наприкінці тріасу — початку юри (190—170 млн. років тому), але викопних залишків найдавніших птахів цього періоду поки не знайдено. У сланцевих піщаниках — відкладеннях мілководної затоки юрського моря (вік близько 150 млн. років) — були знайдені відбитки пір'я й п'ять різної збереженності відбитків кістяка й пір'яний покрив найбільш древньої з відомих нам птахів — археоптерікса — Archaeopteryx lithographica, по розмірах приблизно рівного сороці. Його відносять до особливого підкласу ящерохвостих птахів — Archaeornithes, тому що на відміну від сучасних птахів він мав довгий хвіст приблизно з 20 хребців; до бічних поверхонь кожного хребця кріпилося парне кермове пір'я (мал. 33). Добре розвинене опушення крила, опушене й все тіло. Плече схоже на пташине, пряжка ще не утворилася, а три добре розвинених, вільних пальці кінчаються гострими пазурами. Ключиці срослись у вилочку, а лопатка шаблеподібна; грудина, ще не мала кіля. Задня кінцівка пташиного типу, але із примітивними рисами (розвинена мала гомілкова кістка, не завершене утворення цівки). Як й у багатьох плазунів, в археоптерікса були черевні ребра. Череп рептильного типу, але вже з деякою подобою дзьоба, з витонченими кістками й збільшеними очницями. На верхній і нижній щелепах в альвеолах сидять зуби. Імовірно, археоптерікси були здатні лише перепурхувати з гілки на гілку або до планіруючого польоту, перелітати від дерева до дерева. При переміщеннях у кронах вони, використовували добре розвинені рухливі пальці крил. Незважаючи на багато рис подібності із плазунами, - це, безсумнівно, птах. Археоптерікс представляє примітивну, але спеціалізовану бічну гілку древніх птахів. Предками сучасних птахів, імовірно, були якісь ще більш примітивні ящерохвості птахи.
Рис.
33.
Відбиток
ящерохвостого птаха - археоптерікса

Сучасні й всі інші нині відомі викопні птахи відносяться до підкласу дійсних, або віялохвостих, птахів Neorithes: у них хвостовий відділ хребетного стовпа різко вкорочений, а останні хвостові хребці зливаються в єдину кістку—пігостиль, до якої віялом прикріплюються кермове пір'я. Формується типовий пташиний кістяк крила, на грудині утворюється кіль, завершується утворення цівки, зникають черевні ребра й т.д. Примітивних віялохвостих птахів юрського періоду поки не виявлено, хоча безсумнівно, що в цей період вони вже існували. Найбільш древні залишки віялохвостих птахів виявлені у відкладеннях крейдового періоду (вік приблизно 80—90 млн. років). Їх відносять до двох надрядів. Гесперорніси — Hesperornis sр. були великі, довжиною до метра птахи, що плавали й пірнали, по зовнішньому вигляді нагадували гагар або поганок (мал. 34). Вони втратили здатність до польоту: передні кінцівки й пояс сильно скорочені, а грудина без кіля. Іхтіорнісы — Ichthyornis sр. і інші були розмірами з голуба. Кістяк крила в них типово пташиний, а на грудині добре розвинений
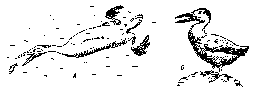
Рис. 34. Гесперорнис (Л) і ихтиорнис (Б) (реконструкція)
кіль; безсумнівно, вони були здатні літати. Від сучасних ці крейдові птахи відрізняються наявністю дрібних зубів на верхній і нижній щелепах і дуже маленьким обсягом мозкової порожнини. Імовірно, це бічні спеціалізовані гілки примітивних віялохвостих птахів. Наприкінці крейди - у третинному періоді кайнозойської ери, приблизно 70-40 млн. років тому, іде інтенсивна адаптивна радіація віялохвостих птахів і виникає більшість сучасних рядів. На жаль, палеонтологічні залишки цього часу нечисленні й не дають уяви про характер і темпи еволюції птахів. Різке збільшення числа видів птахів і формування сучасних рядів за часом збігається з бурхливою еволюцією покритонасінних рослин і комах: зростання потенційних харчових ресурсів сприяло утвореню нових харчових зв'язків і тим самим забезпечувало інтенсивне видоутворення птахів.
Система класу й огляд сучасних груп