

3.8. Определение гетерозиготности в изучаемых группах крупного рогатого скота
По пяти из анализированных генетических маркеров (CSN, PRL, GH/AluI, GH\MspI, PIT-1), в отобранных группах животных, наблюдается преимущественная гомозиготизация в сторону одного из аллелей, приводящая к исчезновению другого. Наиболее гетерогенной группой животных по результатам наших исследований, является частный скот. На фоне отсутствия В-аллеля каппа-казеина, общий уровень гетерозиготности в этой группе по анализируемым маркерам выше, чем в других выборках, за исключение AluI- и MspI-маркеров гена соматотропина (рис. 3.8.1.). По MspI-маркеру в этой группе отмечена самая низкая гетерозиготность из всех опытных групп крупного рогатого скота.
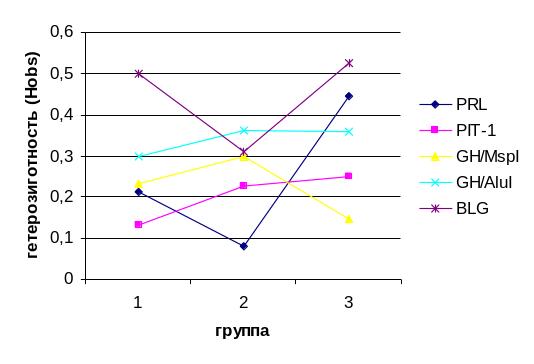
Рис 3.8.1 - Уровень выявленной гетерозиготности (Hobs) в анализируемых группах коров.
1 – черно-пестрая порода, ОАО «Снежка-Госома; 2 – айрширская порода, СПХ Сельцо; 3 - частный скот (Жирятинский р-н, Брянской области).
В литературных источниках имеются сообщения о резком снижении частоты В-аллеля гена каппа-казеина, а в ряде случаев полному его отсутствию у других пород КРС. [Хаертдинов, 1997, Гладырь, 2001, Костюнина, 2004]. В ряде работ приводятся приближенные к установленным нами значениям частот генотипов ВВ гена PRL [Хатами, 2004, Алипанах, 2006]. В некоторых работах отмечается полное отсутствие (-/-) генотипа гена гормона роста и низкая частота VV-генотипа по AluI-маркеру. [Максименко, Хатами, 2004, Lucy, 1993, Kemenes, 1999, Арнаут, 2008]. Преобладание генотипа ВВ локуса PIT-1 отмечается в работе исследователей Cosier с соавторами [Cosier et al., 2008]. Аллель В этого гена широко представлен в группах коров чёрно-пестрой породы в хозяйствах Польши [Dybus, 2004]. Почти одинаковая встречаемость аллелей гена бета-лактоглобулина отмечается в работах Усенбекова и Graml [Усенбекова, 1995, Graml, 1984]. Так же как и в исследуемых нами группах КРС у животных серой украинской породы преобладающим был вариант В β-лактоглобулина [Глазко В. И.] После анализа литературных данных и результатов наших исследований складывается впечатление о существовании общей тенденции по снижению полиморфизма и исчезновению ценных вариантов анализируемых генов.
Косвенно такая картина свидетельствует о гомозиготизации и по аллелям других генов, несущих как положительные, так и патологичные признаки.
В процессе формирования генетической структуры стада по разным причинам участвуют только небольшое число избранных особей, как правило, с неизвестными генотипами. При таком способе селекции возрастает роль генетического дрейфа, из-за которого происходит либо фиксация, либо потеря некоторых хозяйственно ценных аллелей. Потеря аллелей приводит к тому, что генетическая структура стада, вероятно и породы, становится более гомозиготной. Подобная ситуация сложилась в группах коров и телят черно-пестрой породы ОАО "Снежка-Госома", генетические структуры которых резко отличатся. Так, например, в отличие от коров в группе телят утеряны аллель А гена Pit1 и генотип ВВ гена пролактина и произошла еще большая гомозиготизация в сторону (+)-аллеля соматотропина. Использование этих телят в дальнейшей селекционной работе уже не вернет ВВ генотип гена Pit1 и в целом вероятно ускорит темпы обеднения генетической структуры стада.
Кроме того, причинами обеднения генетической структуры стада могут быть:
инбридинг, обусловленный разведением коров в замкнутых популяциях, которое ограничивает приток генов из других популяций и линий [Алтухов Ю.П., 1983; Имашева А.Г., 1998].
искусственный отбор по морфофизиологическим признакам и показателям продуктивности,