

3.7. Анализ сочетания мутаций при определении а и в аллелей гена β-лактоглобулина.
В связи с тем, что в настоящее время разработаны две методики, позволяющие согласованно выявлять А и В варианты гена бета-лактоглобулина, стало возможным предположить наличие рекомбинации между мутациями расположенных в разных частях гена типирующих В-аллель βLG. в основе выбранных нами методик лежит анализ точечных нуклеотидных замен в экзонах 2 и 4. Каждая из этих методик выявляет одну из двух мутаций характерных для В-аллеля. Проводя одновременно анализ по этим методикам можно учитывать эти две замены.
При анализе полиморфизма гена бета-лактоглобулина мы обнаружили несоответствия в определении аллелей этого гена по описанным методикам. На рисунке 3.7.1. представлена электрофореграмма, которая иллюстрирует несовпадения при определении аллельных вариантов по представленным методикам. Особенно интересен результат рестрикции фрагментов экзонов 2 и 4 полученных на ДНК образца 109. При рестрикции фрагмента второго экзона 1247 п.о. эндонуклеазой PvuII выявляется типичная картина характерная для АА – варианта гена бета-лактоглобулина, т.е. отсутствие нуклеотидной замены, обеспечивающей появление дополнительного сайта для PvuII, описанного для ВВ-варианта. Напротив, при рестрикции фрагмента экзона 4 этого же образца эндонуклеазой HaeIII наблюдается появление специфичных для В-аллеля фрагментов.
Частоты несовпадении при определении генотипов по двум вышеописанным методикам в группах коров черно-пестрой, айрширской пород и частного скота составили – 2,7%, 13,3%, 9,7% соответственно.
В таблице 3.7.1. отображены частоты встреченных сочетаний аллельных вариантов при определении полиморфизма гена βLG по методикам Гладырь и Medrano [Гладырь и др., 2001 и Medrano, et al., 1990]. С наивысшей частотой встречались сочетания АВ/ВВ (0,61) и АА/ВВ (0,22). Сочетание ВВ/АВ было самым редким, а сочетания АВ/АА и ВВ/АА не обнаружены. Генотипы АВ/АВ неотличимы от обычных гетерозигот АВ.
Таблица 3.7.1
Частоты сочетаний мутаций в генотипах КРС при определении полиморфизма гена бета-лактоглобулина по описанным методикам.
Варианты гена βLG по методике Гладырь и др., 2001 г. |
Варианты гена βLG по методике Medrano, et al.,1990
|
|||
|
ВВ |
АВ |
АА |
|
АВ |
0.61 |
|
- |
|
АА |
0.22 |
0.11 |
|
|
ВВ |
|
0.055 |
- |
|
На рисунке 3.7.2 представлена схема, раскрывающая филогенетические связи между разными вариантами гена бета-лактоглобулина [Formaggioni P., 1999]. В основе филогенетического дерева авторы расположили В-аллель βLG. Добавляя к этому варианту определенные мутации можно прийти ко всем остальным аллелям.
Проведенные нами исследования по выявлению двух мутаций определяющих В-вариант в целом соответствуют приведенной филогенетической связи А- и В-аллелей, но выявленные нами сочетания мутаций не вкладываются в эту схему. Руководствуясь результатами наших исследований, можно предположить, что существуют промежуточные между А- и В-аллелями варианты или, что произошла рекомбинация между мутациями во втором и четвертом экзонах гена бета-лактоглобулина с образованием новых аллелей (Рис. 3.7.3.).
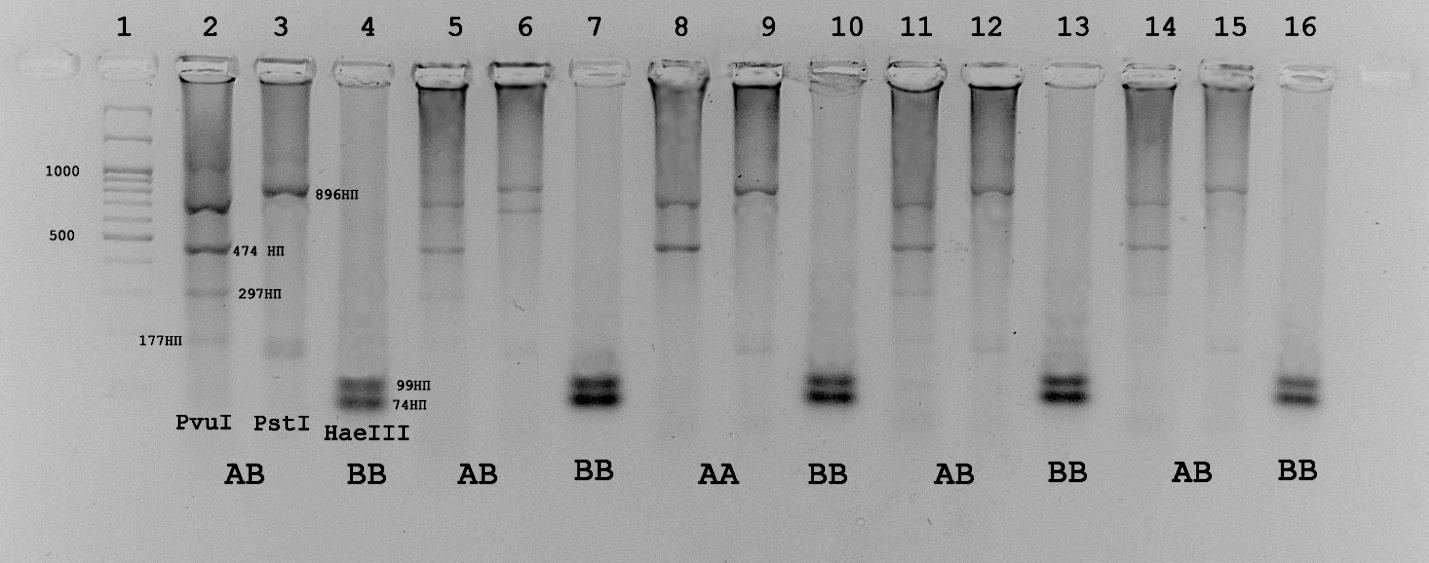
Рис.3.7.1 - Электрофореграмма продуктов рестрикции при определении вариантов гена бета-лактоглобулина по методикам Гладырь и др., 2001 и Medrano, et al., 1990. 1 – маркер молекулярных масс М27(СибЭнзим). 2,3,4-анализ образца 23; 5, 6, 7 – образец 99; 8, 9, 10 – образец 109; 11, 12, 13 - образец 96; 14, 15, 16 – образец 181. Каждый первый из тройки трек – рестрикция эндонуклеазой PvuII, каждый второй - продукты рестрикции PstI, каждый третий - продукты рестрикции HaeIII .
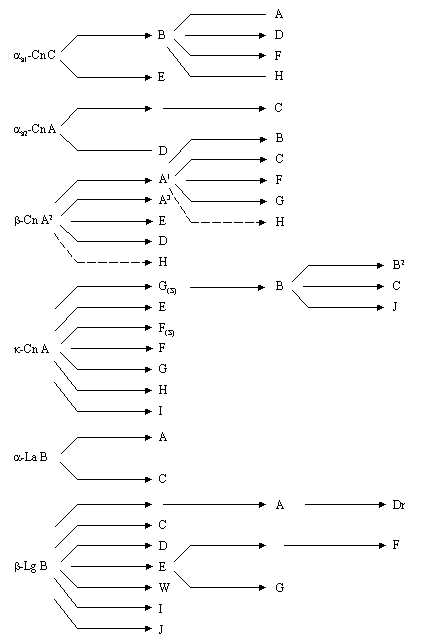
Рис. 3.7.2 - Схема происхождения генетических вариантов гена бета-лактоглобулина гена бета-лактоглобулина (по P. Formaggioni et all.,1999)

Рис. 3.7.3 - Вероятное положение промежуточных вариантов между А- и В-аллелями в схеме филогении полиморфизма гена бета-лактоглобулина.