

3.6. Исследование генетической структуры групп крупного рогатого скота по гену бета-лактоглобулина (βLg).
Идентификацию аллельного полиморфизма βLG по точечной замене в нуклеотидной последовательности в экзоне 4 предложил Medrano, et al. 1990 год. Эта мутация предполагает появление дополнительного сайта рестрикции для эндонуклеазы HaeIII и замене в пептидной последовательности аминокислоты Val на Ala для В варианта в положении 118.
Другая методика, определения аллелей А, В была разработана Гладырь [Гладырь и др.,2001]. По этой методике вариант В определяется по мутации второго основания триплета GAT>GGT во втором экзоне, приводящего к замене аминокислоты в положении 64.
По методике Medrano, et al., 1990 амплифицировали фрагмент ДНК включающий участок 4 интрона и 4 экзона 247 п.о. Вариант В от А отличается наличием дополнительного сайта для рестриктазы HaeIII. После обработки продуктов ПЦР этой рестриктазой появляются специфичные для этих аллелей фрагменты: 148 и 99п.о. – для А варианта и 99, 74 и74 п.о. для В варианта. На рисунке 3.6.1. показана электрофореграмма фрагментов ДНК после рестрикции эндонуклеазой HaeIII.
По методике (Гладырь и др., 2001) амплифицировали фрагмент длиной 1248 п.о. После рестрикции по сайтам PvuII наблюдали появление специфичных фрагментов длиной 774 и 474 п.о. соответствующих аллелю А. Точковая мутация в нуклеотидной последовательности варианта В обуславливает образование дополнительно сайта рестрикции PvuII и приводит к появлению трёх специфичных для аллеля В фрагментов длиной 774, 297 и 177 п.о. Фрагменты с длинами соответствующие аллелю D выявлены не были. Рестрикция PstI позволяет определять аллель С. В анализируемых образцах вариант C также обнаружен не был. Пример определения варианта гена бета-лактоглобулина по этой методике приведен на рисунке 3.6.2.
Выявление аллельных вариантов гена βLG мы проводили по двум выше представленным методикам, в основе которых лежит анализ точечных нуклеотидных замен в экзонах 2 и 4.
При анализе полиморфизма бета-лактоглобулина с разной частотой были выявлены три генотипа. Результаты анализа генетической структуры изучаемых групп КРС и частоты несовпадений в определении генотипов по описанным методикам указаны в таблице 3.6.1. При оценке полиморфизма гена бета-лактоглобулина в исследуемых группах КРС в разной степени наблюдается преобладание В-аллеля, максимальная частота этого аллеля зафиксирована в группе коров айрширской породы, там она достигает 77%. В этой же группе отмечена наивысшая частота АА-генотипа. Иная картина наблюдается в группах коров черно-пестрой породы и частного скота. В этих группах чаше всего встречались животные с гетерозиготным генотипом. В выборке частного скота отмечены приближенные значения частот А – и В – генотипов. Наглядно соотношение частот генотипов гена β – лактоглобулина в исследуемых группах КРС представлено на рисунке 3.6.3.
Во всех анализируемых группах крупного рогатого скота выявленные частоты аллелей достоверно не отклонятся от теоретически рассчитанных.
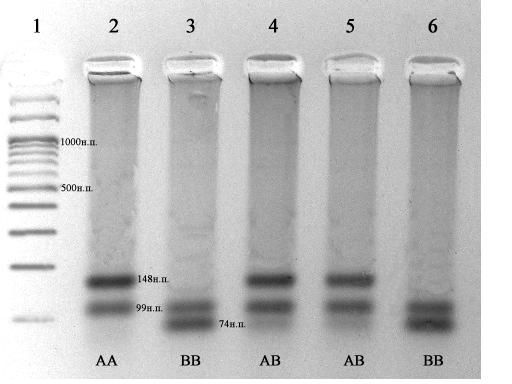
Рис. 3.6.1 - Электрофоретический анализ продуктов рестрикции, при определении генотипа β – лактоглобулина по методике Medrano, et al., 1990. 1 – маркер молекулярных масс М27 (СибЭнзим); Генотипы образцов 2-6 указаны на фотографии
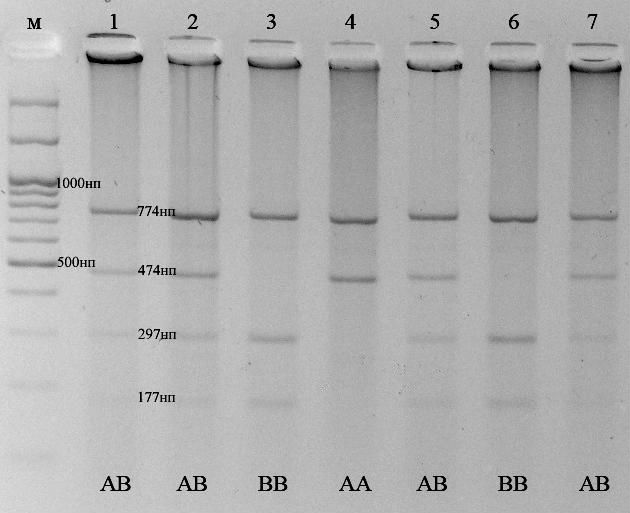
Рис 3.6.2 - Электрофоретический анализ продуктов рестрикции, при определении генотипа β – лактоглобулина по методике Гладырь и др., 2001. 1 – маркер молекулярных масс М27 (СибЭнзим); Генотипы образцов 2-7 указаны на фотографии
Таблица 3.6.1
Генетическая структура анализируемых групп КРС по гену бета-лактоглобулина определенная по методикам Гладырь и др., 2001 и Medrano, et al., 1900
порода |
n |
Частота аллелей и генотипов, % по методике Гладырь и др.,2001 |
Частота аллелей и генотипов, % по методике Medrano, et al., 1900 |
Частота несовпадений генотипов, % |
||||||||||||
АA |
AВ |
ВB |
А |
В |
АA |
AВ |
ВВ |
А |
В |
|||||||
Черно-пестрая |
55 |
19,4 |
50 |
30,5 |
0,41 |
0,58 |
19,4 |
47,2 |
33,3 |
0,43 |
0,57 |
2,7 |
||||
Hex |
0,493 |
0,49 |
|
|||||||||||||
Hobs |
0,5 |
0,472 |
|
|||||||||||||
χ2 |
0,0043 |
|
|
|||||||||||||
Айрширская |
50 |
6,6 |
31,3 |
62,2 |
0,22 |
0,77 |
2,2 |
26,6 |
71,1 |
0,15 |
0,84 |
13,3 |
||||
Hex |
0,345 |
0,262 |
|
|||||||||||||
Hobs |
0,311 |
0,266 |
|
|||||||||||||
χ2 |
0,8445 |
- |
|
|||||||||||||
Частный скот |
146 |
21,3 |
52,4 |
26,2 |
0,47 |
0,52 |
18,0 |
42,6 |
39,3 |
0,39 |
0,6 |
9,7 |
||||
Hex |
0,498 |
0,477 |
|
|||||||||||||
Hobs |
0,524 |
0,426 |
|
|||||||||||||
χ2 |
0,3963 |
- |
|
|||||||||||||

Рис.3.6.3- Соотношение частот генотипов β-лактоглобулина в анализируемых группах КРС. 1 – группа коров черно-пестрой породы, ОАО «Снежка-Госома; 2 – частный скот (Жирятинский р-н, Брянской области); 3 – группа коров айрширской породы, СПХ Сельцо.
В таблице 3.6.2 приведены частоты А-аллеля β-лактоглобулина в группах крупного рогатого скота разных пород. Из таблицы видно, что преобладание В-аллеля наблюдается в разных регионах мира. Исключением являются некоторые группы коров голштинской и чешской красной. В этих группах частота А-аллеля превышает частоту В-аллеля. Кроме того, для разных групп крупного рогатого скота голштинской породы из разных регионов мира в литературе приводятся близкие значения частот аллелей. Так, например, в группе голштинов из США, Ирана, Мексики, Украины, Аргентины частоты А-аллеля составляют - 0,530, 0529, 0,470, 0460, 0,43 соответственно. Вероятно, что для голштинской породы частота аллелей гена бета-лактоглобулина может является породоспецифичной характеристикой. Ранее, Глазко ранее отмечал у голштинов по локусу каппа-казеина незначительные межгрупповые различия, не зависящие от региона разведения [Глазко, 2006].
Частота А-аллеля в исследуемых нами группах частного скота и коров черно-пестрой породы приближенно соответствует частотам этого аллеля в разных группах КРС голштинской породы, что косвенно может быть связано с голштинизацией.
Таблица 3.6.2
Частота А-аллеля бета-лактоглобулина в группах КРС Брянской области и в других регионах
порода |
Частота А-аллеля |
Автор |
Регион |
Чешская красная |
0,571 |
M Zatoń-Dobrowolskaet al., 2006 |
Польша |
Caracu |
0,57 |
Kemenes et al.1999 |
Бразилия |
Голштинская |
0,530 |
A. Barreras et al., 2001 |
США |
Голштинская |
0,529 |
Heidari et al. |
Иран |
Якутская |
0,516 |
Гладырь и др., 2000 |
Россия |
Черно-пестрая |
0,48 |
Гладырь и др., 2000 |
Россия |
Частный скот |
0,47 |
Собственные данные |
Россия, Брянск |
Голштинская |
0,470 |
А. Barreras et al., 2002 |
Мексика |
Голштинская |
0,460 |
Копылова К.В., и др., 2009 |
Украина |
Romanian Spotted |
0,450 |
Ilie D et al., 2008 |
Румыния |
Голштинская |
0,43 |
A. G. Bonvillani et al., 1998 |
Аргентина |
Черно-пестрая |
0,41 |
Собственные данные |
Россия, Брянск |
Швицкая |
0,406 |
Гладырь и др., 2000 |
Россия |
Nelore (Bos indicus) |
0,40 |
Kemenes et al.1999 |
Бразилия |
Польская красная |
0,397 |
M Zatoń-Dobrowolskaet al., 2006 |
Польша |
Tharparkar(Bos indicus) |
0,39 |
S. Rachagani et al., 2006 |
Индия |
Gyr |
0,37 |
Kemenes et al.1999 |
Бразилия |
Brown of Maramures |
0,333 |
Ilie D et al., 2008 |
Румыния |
Эстонская голштинская |
0,319 |
Sirje Varv, et al., 2009
|
Эстония |
Bubalus bubalis |
0,301 |
Rajesh K et al., 2007 |
Индия |
Jersey × Zebu, |
0,30 |
Rajesh K. PATEL et al., 2007 |
Индия |
Украинская черно-пестрая |
0,278 |
Малиенко В.А., и др., 2008
|
Украина |
Голштино-фризская × Zebu |
0,26 |
Rajesh K. PATEL et al., 2007 |
Индия |
Красная горбатовская |
0,246 |
Гладырь и др., 2000 |
Россия |
Айрширская |
0,22 |
Собственные данные |
Россия, Брянск |
Hanwoo |
0,208 |
Kim, J et al., 1998 |
Корея |
Германская красная |
0,191 |
M Zatoń-Dobrowolskaet al., 2006 |
Польша |
Эстонская |
0,188 |
Sirje Varv, et al., 2009 |
Эстония |
Sahiwal (Bos indicus) |
0,17 |
S. Rachagani et al., 2006 |
Индия |
Украинская черно-пестрая |
0,165 |
Копылова К.В., и др., 2009 |
Украина |
Корейская |
0,153 |
Chung, E. R et al., 1998 |
Корея |
Эстонская красная |
0,138 |
Sirje Varv, et al., 2009
|
Эстония |
Датская красная |
0,110 |
M Zatoń-Dobrowolskaet al., 2006 |
Польша |
Абердин-ангусская |
0,2 |
Копылова К.В., и др., 2009 |
Украина |