

3.5. Сравнение распространения аллелей гена соматотропина по MspI- и AluI – маркерам у крс в Брянской области и других регионах.
В литературе отмечена крайне низкая частота Msp(-)-аллеля у северо-европейских пород КРС. Наивысшая частота этого аллеля наблюдается у зебу [Lagziel et al., 2000]. Возможно, это является одной из причин высокой жирности молока зебу. Эта особенность сделала зебу ценным материалом для гибридизации и выведения новых жирномолочных пород крупного рогатого скота. Жирномолочность гибридных коров, обусловленная наследственным влиянием зебу, оказалась наивысшей и составила 4,15 %, что на 0,58–0,32 % больше, чем у чистопородных сверстниц [Караев Г. С., 2009]. Преобладание частоты Msp(-)-аллеля у зебу, может свидетельствовать о зебувидном происхождении этого аллеля [Keiko K. de M. et al., 2004]
Анализ литературных данных по частотам MspI(-)-аллеля, приведенных в таблице 3.5.1 позволяет условно выделить три группы.
В первую группу в основном входят европейские и североевропейские породы КРС, у них частота этого аллеля колеблется от 0,02 до 0,19. В эту же группу вошли анализированные нами стадо коров айрширской породы и частный беспородный скот.
Вторую группу составляют породы со средним значением часты MspI(-)-аллеля. В основном это породы украинской и русской селекции. Частота (-) аллеля в этой группе колеблется от 0,25 до 0,38. В эту группу попала проанализированная нами выборка коров черно-пестрой породы.
Особую группу составляют зебу и монгольский як. Частота MspI(-)-аллеля в ней достигает 0,94.
При сопоставлении частот Msp(-)-аллеля в стадах КРС одной породы отмечается значительный уровень внутрипородной изменчивости, например: чёрно-пёстрая – Брянск (0,26) и чёрно-пёстрая – Москва (0,13); немецкая чёрно-пёстрая – Германия, Думмерсторф (0,08) и немецкая чёрно-пёстрая – Германия (0,18). Такая ситуация, вероятно, возникла в результате изоляции и разнонаправленного селекционного давления внутри отдельных стад.
Таблица 3.5.1
Сравнение частот аллеля (-) и V - аллелей гена соматотропина по MspI-и AluI-маркерам.
группа |
Порода |
MspI(-) |
AluI(V) - |
Ссылки |
I |
Голштинская |
0,02 |
0,08 |
Lagziel et al., 2000 |
Ярославская |
0,02 |
0,56 |
Хатами С.Р., 2004 |
|
Braunvieh коричневая |
0,02 |
|
Mitra A., et al, 1994 |
|
Норвежская красная |
0,05 |
|
Hoj et al., 1993 |
|
Симментальская |
0,06 |
0,56 |
Lagziel et al., 2000 |
|
Норвежская красная |
0,07 |
|
Lagziel et al., 2000 |
|
Немецкая чёрно-пестрая |
0,08 |
0,131 |
Хатами С.Р., 2004 |
|
Красная горбатовская |
0,09 |
|
Туркова, 2003 |
|
Чёрно-пёстрая |
0,13 |
0,38 |
Хатами С.Р., 2004 |
|
Частный скот, Брянск |
0,13 |
0,225 |
Собственные данные |
|
Польская чёрно-пёстрая |
0,145 |
|
Dubis A., 2002 |
|
Джерсейская |
0,17 |
|
Lagziel et al., 2000 |
|
Айрширская, Брянск |
0,17 |
0,265 |
Собственные данные |
|
Немецкая черно-пестрая |
0,18 |
0,2 |
Mitra A., et al, 1994 |
|
Датская красная |
0,19 |
|
Hoj et al., 1993 |
|
II |
Карпатская коричневая |
0,25 |
|
Lagziel et al., 2000 |
Чёрно-пёстрая (Брянск) |
0,26 |
0,145 |
Собственные данные |
|
Бурая карпатская |
0,26 |
|
Глазко В.И., |
|
Pesisir (Индонезия) |
0,28 |
0,16 |
SUTARNO, 2010 |
|
Монгольская |
0,3 |
|
Туркова, 2003 |
|
Brangus bulls |
0,31 |
0,4 |
M.G. Thomas, 2007 |
|
Madura |
0,315 |
0 |
SUTARNO, 2010 |
|
Серая украинская |
0,32 |
|
Lagziel et al., 2000 |
|
Серая украинская |
0,326 |
|
Глазко В.И., |
|
Бестужевская (Уфа) |
0,38 |
|
Ильясов А.Г., 2008 |
|
III |
Sahywal (зебу) |
0,6 |
0,04 |
Mitra A., et al, 1995 |
Гир (зебу) Бразилия |
0,81 |
0 |
Keiko Kusamura de Mattos,, 2004 |
|
Гир (зебу) |
0,94 |
0,04 |
Lagziel et al., 2000 |
|
Монгольский як |
0,94 |
0 |
Туркова, 2003 |
|
|
Онгол (зебу) |
0,94 |
0 |
Lagziel et al., 2000 |
При сопоставлении частот аллелей MspI(-)- и V-аллелей определяемых двумя мутациями расположенными в разных частях гена, было обнаружено, что в породах с максимальной частотой MspI(-)-аллеля частота V-аллелей минимальна и наоборот. Так в стаде зебувидного скота с частотой MspI(-)-аллеля 0,94 аллель V отсутствует, а у ярославской породы при частоте MspI(-)-аллеля – 0,02, частота V-аллель соствляет 0,56). Эта зависимость подтвердилась при корреляционном анализе (коэф. коррел.=-0,65).
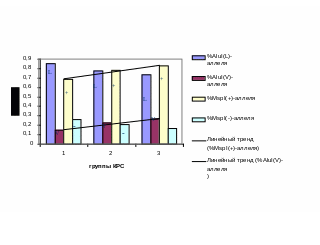
Рис. 3.5.1 - Соотношение частот аллелей соматотропина по MspI- и AluI-маркерам в анализированных группах КРС. 1 – группа коров черно-пестрой породы, ОАО «Снежка-Госома; 2 – частный скот (Жирятинский р-н, Брянской области); 3 – группа коров айрширской породы, СПХ Сельцо.
Аналогичная зависимость прослеживается при сравнении частот (-) и V - аллелей в трёх анализируемых нами группах КРС, значение коэффициента корреляции составило -0,80. На рисунке 3.5.1 приведена гистограмма с соотношением аллелей гормона роста по обоим маркерам в анализируемых группах КРС. Линии тренда проходящие по вершинам столбцов с частотами (+) и V-аллелей в анализируемых группах КРС практически параллельны.
Таким образом, при отборе Msp(-)-аллеля, с которым ассоциированы высокая жирность молока, в стаде может происходить обеднение V-аллелем, также благоприятным с хозяйственной точки зрения и наоборот.