

3.2. Генетическая структура исследуемых групп крупного рогатого скота по гену пролактина (prl).
На всех проанализированных образцах ДНК был амплифицирован фрагмент длинной 156 н.п. После обработки рестриктазой RsaI выявляли генотипы животных: генотипу АА соответствовал фрагмент 156 н п., генотипу АВ – 156, 82 и 72 н.п., генотипу ВВ – 82 и 74 н.п.
На рисунке 3.2.1. представлена электрофореграмма с продуктами рестрикции при определении генотипов пролактина.
При тестирование животных изучаемых групп, были выявлены А- и В-аллели гена пролактина.
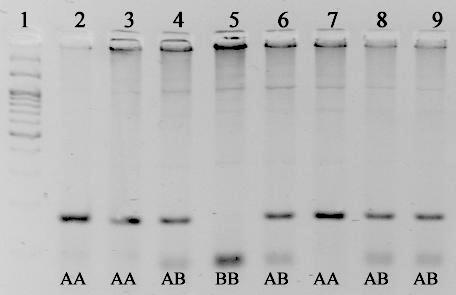
Рис. 3.2.1 Электрофоретический анализ продуктов рестрикции, при определении генотипа пролактина. 1 – маркер молекулярных масс М27 (СибЭнзим); Генотипы образцов указаны на фотографии
Из таблицы 3.2.1. видно, что общим для анализируемых групп животных является превышение частоты А-аллеля над частой В-аллеля. Максимальная частота А-аллеля зафиксирована в группе коров айрширской породы (0,94).
Таблица 3.2.1
Генетическая структура анализируемых групп коров по гену пролактина
Группы животных |
n |
Частота встречаемости генотипов |
χ2 |
Частота встречаемостиаллелей |
Hobs |
Hex |
|||
АА |
АВ |
ВВ |
А |
В |
|||||
Коровы черно-пестрой породы |
55 |
0,75 |
0,21 |
0,04 |
0,754 |
0,87 |
0,13 |
0,2115 |
0,2188 |
Телята черно-пестрой породы |
16 |
0,71 |
0,28 |
0 |
0,119 |
0,84 |
0,15 |
0,2857 |
0,2449 |
Коровы айрширской породы |
50 |
0,87 |
0,12 |
0 |
1,290 |
0,94 |
0,06 |
0,0816 |
0,115 |
Частный скот |
146 |
0,39 |
0,55 |
0,06 |
7,243 |
0,66 |
0,33 |
0,5455 |
0,4444 |
P=0,05
При анализе частот генотипов телят и коров черно-пестрой породы, содержащихся в одном хозяйстве отмечено, что в группе телят в отличие от группы коров отсутствует ВВ-генотип. Это, вероятно, указывает на высокую внутривидовую изменчивость частот аллей и быстрое изменение генетической структуры стада, даже внутри одного хозяйства. В этих группах выявлены приближенные значения наблюдаемой и ожидаемой гетерозиготности.
В группе коров айрширской породы В-аллель обнаружен только в составе гетерозигот, которые в группе представлены с низкой частотой (12%). Такое распределение частот, вероятно, является следствием гомозиготизации преимущественно по одному аллелю и может привести к потери варианта гена.
В группе частного скота частота В-аллея максимальна из всех опытных групп (33%). Внутри группы чаще всего встречаются животные с гетерозиготным генотипом, их частота составляет 55%, за счет этого доля В-аллеля в этой группе максимальна (33%), несмотря на то, что частота ВВ-генотипа незначительно отличается от частоты этого генотипа в других группах.
В изучаемых группах КРС не вывалено достоверного отклонения частот аллей от теоретически ожидаемого,
И з
рисунка 3.2.2 видно, что в стадах КРС
содержащихся в условиях специализированных
хозяйств, наблюдается высокая частота
гомозиготного генотипа АА. В группе
частного скота наблюдается преимущественное
присутствие гетерозигот, что отразилось
на значении наблюдаемой гетерозиготности.
з
рисунка 3.2.2 видно, что в стадах КРС
содержащихся в условиях специализированных
хозяйств, наблюдается высокая частота
гомозиготного генотипа АА. В группе
частного скота наблюдается преимущественное
присутствие гетерозигот, что отразилось
на значении наблюдаемой гетерозиготности.
Рис.3.2.2. Соотношение частот генотипов пролактина в анализированных группах КРС. 1 – группа коров черно-пестрой породы, ОАО «Снежка-Госома; 2 – частный скот (Жирятинский р-н, Брянской области); 3 – группа коров айрширской породы, СПХ Сельцо.
В таблице 3.2.2 представлены частоты В-аллеля пролактина выявленные группах крупного рогатого скота разных пород. Из таблицы видно, что в основном А-аллель является доминирующей во многих регионах мира. В анализируемых нами группах коров черно-пестрой породы и частного скота частота В-аллеля приближенно соответствует среднемировым значениям. В группе коров айрширской породы отмечена одна из самых низких зафиксированных частот этого аллеля (0,06). При анализе данных таблицы 3.2.2. можно установить, что частота аллелей пролактина не является породоспецифичной. Как и в случае с полиморфизмом гена каппа-казеина, наблюдается широкий разброс частот внутри породы КРС, даже в одном регионе. Это наблюдается как в стадах B. taurus, так и B. indicus.
Таблица 3.2.2
Частоты В-аллеля пролактина в группах КРС Брянской области и других регионов.
Порода |
Частота В-аллеля |
Ссылка |
Регион |
Jersey cattle |
0,691 |
A Dybus et al., 2005 |
Польша |
Red Sindhi (Zebu) |
0,54 |
A. Ratna Kumari et al., 2008 |
Индия |
Kankrej |
0,491 |
Sacravarty G. et al., 2008 |
Индия |
Najdi Cattle |
0,429 |
S. Sharifi et al., 2010 |
- |
Голштино-фризская |
0,417 |
K. Wojadak-Maksymiec et al., 2008 |
Польша |
Gir |
0,39 |
A. Ratna Kumari et al., 2008 |
Индия |
Черно-пестрая |
0,39 |
Хатами С.Р., |
Германия |
Ярославская |
0,35 |
Хатами С.Р., |
Россия, Ярославская обл. |
Частный скот |
0,33 |
Собственные данные |
Россия, Брянск |
Корейская |
0,322 |
Chung, E. R et al., 1998 |
Корея |
Slovak Pinzgauer |
0,32 |
P. Chrenek et al., 1988 |
Словакия |
Черно-пестрая |
0,29 |
Масуд. А. И др., 2007 |
Россия |
Черно-пестрая |
0,23 |
Собственные данные |
Россия, Брянск |
Красно-пестрая |
0,21 |
Масуд. А. И др., 2007 |
Россия |
Литовская черно-пестрая |
0,21 |
Rūta Skinkytė et al., 2005 |
Литва |
Черно-пестрая |
0,146 |
A Dybus et al., 2005 |
Польша |
Айрширская |
0,14 |
Юдина и др., 2001 |
Россия |
Черно-пестрая |
0,138 |
A Dybus et al., 2002 |
Польша |
Литовская красная |
0,13 |
Rūta Skinkytė et al., 2005 |
Литва |
Slovak Pied |
0,13 |
P. Chrenek et al., 1988 |
Словакия |
Sahiwal (Zebu) |
0,12 |
A. Ratna Kumari et al., 2008 |
Индия |
Montebeliard |
0,11 |
N Ghasemi et al., 2009 |
Иран |
Ongole |
0,1 |
A. Ratna Kumari et al., 2008 |
Индия |
Красная горбатовская |
0,08 |
Юдина и др., 2001 |
Россия |
Айрширская |
0,06 |
Собственные данные |
Россия, Брянск |
Голштинская |
0,05 |
P. Chrenek et al., 1988 |
Словакия |
Черно-пестрая |
0,046 |
Хатами С.Р., |
Россия, Московская обл. |