

2.8. Анализ полиморфизма гена гипофизарного фактора транскрипции (pit-1)
Для выявления полиморфизма гена гипофизарного фактора транскрипции, с помощью праймеров:
PIT-1 /HinfI: 5/- CAA-TGA-GAA-AGT-TGG-TGC-3/
PIT-1 /HinfI: 5/- TCT-GCA-TTC-GAG-ATG-CTC -3/ [Moody, 1995] амплифицировали фрагмент ДНК длиной 1355 п.о.
ДНК денатурировали при 94°С в течение 4 минут далее проводили 35 циклов амплификации в следующем режиме: 94°С – 1минут, отжиг праймеров при 54°С - 1 минут элонгация при 72°С – 1минут Конечный этап синтеза проводили при 72°С в течение 4 минут.
Для определения полиморфизма гена соматотропина 3 мкл ПЦР смеси обрабатывали 5 ед. эндонуклы рестрикции HinfI в 1×буфере «O» фирмы СибЭнзим (Россия) при 37 0С течение 1 часа.
Появление фрагментов – 660 н.п., 425 н.п. и 270 н.п. после обработки продуктов ПЦР рестриктазой соответствует (А) - аллелю гена; фрагменты - 660 н.п., 385 н.п. и 270 н.п. указывает на (В)-вариант.
Для визуализации фрагментов ДНК проводили горизонтальный электрофорез при 10 В/см в 1×ТВЕ буфере на 1,0 % агарозной пластине содержащей 0,1 мкг/мл этидия бромида.
2.9. Статистическая обработка результатов
Частоту встречаемости генотипов определяли по формуле:
р = n/N, где р – частота генотипа, n – количество особей,
имеющих определенный генотип, N – число особей.
Частоту отдельных аллелей определяли по формуле:
РА = (2nAA+nAB) : 2N и qB = (2nBB+nAB) : 2N, где PА – частота аллеля А, qB – частота аллеля В, N – общее число аллелей.
Для оценки достоверности отклонения распределения выявленных частот аллелей от теоретически ожидаемого использовали критерий хи-кадрат.
Статистическую обработку проводили с помощью программ «Statistika 6.0.», «Popgene 1.31.», MS Excel 2003 (Microsoft) и «Pop Stat», разработанной Гусаревой Е. и В.Рюхко, 2001.
3. Результаты и обсуждение
3.1. Генетическая структура исследуемых групп крупного рогатого скота по гену каппа-казеина (csn3)
Нами определены генотипы по гену каппа-казеину 55 коров и 16 телят черно-пёстрой породы ОАО "Снежка-Госома", аирширской породы (n=50) СПХ Сельцо и 146 частных коров на территории Жирятинского района Брянской области.
Аллельные варианты А и В белка каппа-казеина отличаются двумя аминокислотными заменами (Thr 136Iso и Asp 148Ala) и кодируются разными аллелями гена.
На рисунке 3.1.1. представлена электрофореграмма с примером определения варианта гена каппа-казеина с помощью рестриктазы HinfI.
Проведенный нами анализ показал отсутствие полиморфизма локуса каппа-казеина. Во всех анализируемых группах был отмечен только генотип АА.
В таблице 3.1.1. приведены частоты В-аллеля CSN3 в группах КРС разных пород. Из таблицы видно, что частоты аллелей каппа-казеина в разных стадах одной породы различны. Так, например, внутри самой распространенной черно-пестрой породы КРС частота В-аллеля колеблется от 0,34 (Краснодарский край) до полного отсутствия (Брянск) или внутри симментальской породы – 0,560 (Словакия) и 0,16 (Рязанская обл.). Эти данные указывают на то, что частота аллельных вариантов гена каппа-казеина не является породоспецифичной характеристикой. Вероятно, распределение частот аллелей этого гена, определяется особенностями воспроизводства стада, в процессе которого не учитывается генотип производителей.
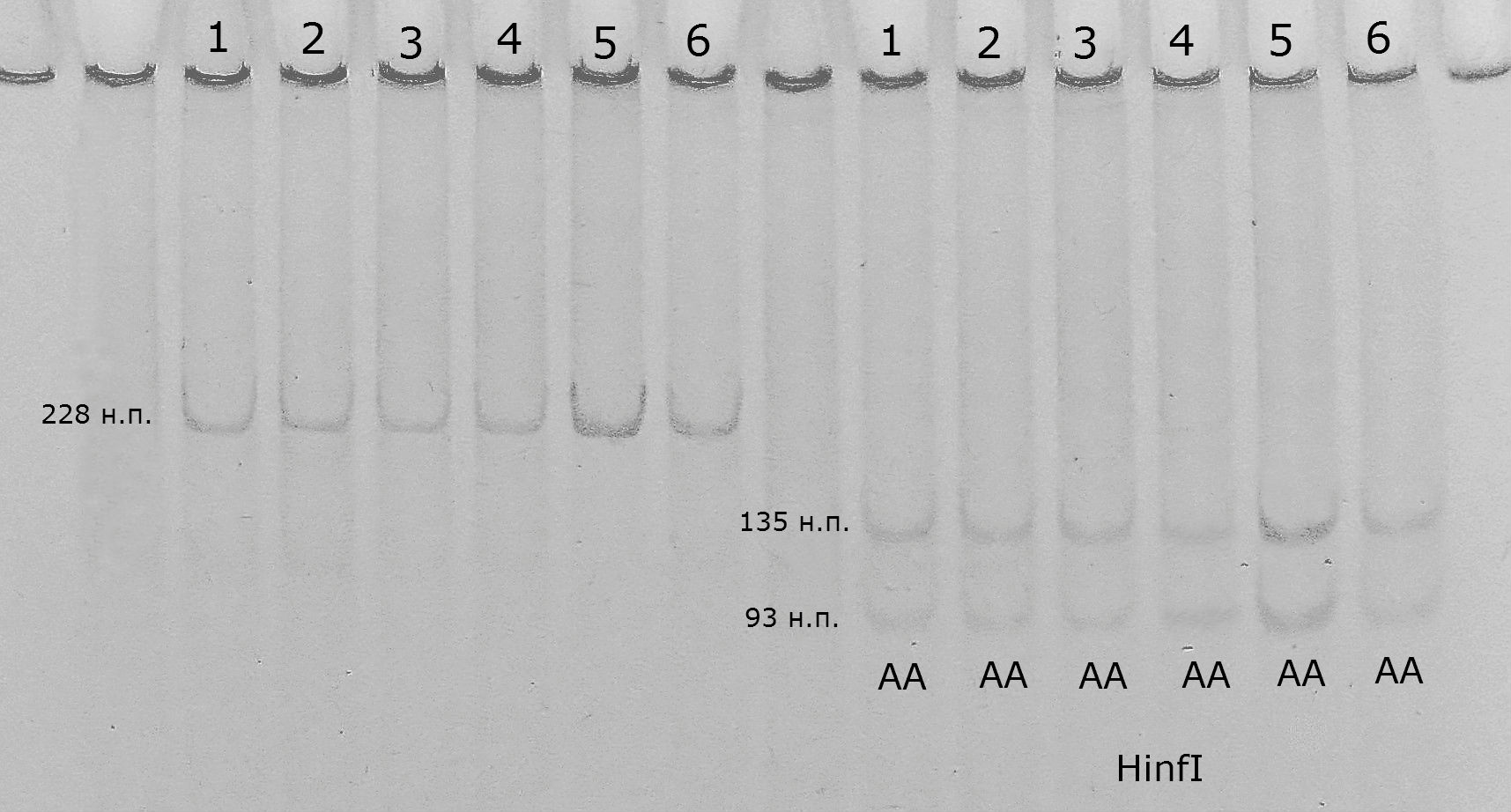
Рис. 3.1.1 - Электрофоретический анализ продуктов рестрикции, при определении генотипа каппа-казеина; слева на право – 1-6 продукты ПЦР; 1-6 продукты рестрикции HinfI. Генотипы образцов указаны на фотографии.
Таблица 3.1.1
Частоты В-аллеля каппа-казеина в группах КРС Брянской области и других регионов.
Порода |
Частота В-аллеля |
Ссылка |
Регион |
Симментальская |
0,560 |
D. Vasicek et al., 1995 |
Словакия |
Ярославская |
0,529 |
Журавель Е.В., 1999 |
Россия |
Серая украинская |
0,500 |
Журавель Е.В., 1999 |
- |
Бурая карпатская порода |
0,50 |
Копылова К. В., 2006 |
Украина |
Ярославская |
0,475 |
Сулимова Г.Е. и др., 2007 |
Россия, Ярославская обл. |
Симментальская |
0,470 |
A. Felenczak et al.,2006 |
Польша |
Ярославская |
0,44 |
Хабибрахманова Я.А., 2009 |
Россия, Ярославская обл. |
Красно-пестрая |
0,436 |
Баршинова А.В., и др., 2004 |
Россия, Липецкая обл. |
Голштинская |
0,410 |
Усенбеков Е.С.,1995 |
Россия, Ленинградская обл. |
Черная уэльская |
0,397 |
Журавель Е.В., 1999 |
Англия |
Красная польская |
0,380 |
Копылова К. В., 2006 |
Украина |
Черно-пестрая |
0,380 |
Усенбеков Е.С.,1995 |
Россия, Ленинградская обл. |
Бурая карпатская |
0,364 |
Журавель Е.В., 1999 |
- |
Аргентинская голштинская |
0,353 |
C.D. Golijow et al.,1999 |
Аргентина |
Аргентинская креольская |
0,344 |
C.D. Golijow et al.,1999 |
Аргентина |
Черно-пестрая |
0,34 |
Денисенко Е.А., 2004 |
Россия, Краснодарский край |
Красно-пестрая |
0,33 |
Юхманова Н.А., 2004 |
Мордовия |
Калмыцкая |
0,324 |
Сулимова Г.Е. и др., 2007 |
Калмыкия |
Бестужевская |
0,293 |
Сулимова Г.Е. и др., 2007 |
Татарстан |
Красно-пестрая |
0,27 |
Алипамах М., 2006 |
Россия, Воронежская обл. |
Якутская |
0,253 |
Сулимова Г.Е. и др., 2007 |
Якутия |
Симментальская |
0,245 |
Коновалова Е.Н. и др., 2004 |
Россия, Орловская обл. |
коровы татарского типа |
0,17-0,27 |
Ахметов Т.М., 2009 |
Татарстан |
Холмогорская |
0,23 |
Глотова Г.Н., 2007 |
Россия, Рязанская обл. |
Чёрно-пёстрая (голшт.) |
0,22 |
Хабибрахманова Я.А., 2009 |
Россия, Московская обл. |
Черно-пестрая |
0,22 |
D. Vasicek et al., 1995 |
Словакия |
Голштино-фризская |
0,21 |
D. Vasicek et al., 1995 |
Словакия |
Холмогорская (голшт.) |
0,20 |
Хабибрахманова Я.А., 2009 |
Россия, Московская обл. |
Черно-пестрая |
0,170 |
Яцына О.А., 2010 |
Беларусь |
Черно-пестрая |
0,17 |
Алипамах М., 2006 |
Россия, Московская обл. |
Симментальская (голшт.) |
0,16 |
Хабибрахманова Я.А., 2009 |
Россия, Рязанская обл. |
Черно-пестрая |
0,153 |
Сулимова Г.Е. и др., 2007 |
Россия, Московская обл. |
Абердин-ангусская |
0,150 |
Арнаут Е.А., 2008 |
Украина |
Украинская красно-пестрая |
0,105 |
Арнаут Е.А., 2008 |
Украина |
Украинская черно-пестрая |
0,099 |
Малиенко В.А. и др., 2008 |
Украина |
Черно-пестрая |
0 |
Собственные данные |
Россия, Брянск |
Айрширская |
0 |
Собственные данные |
Россия, Брянск |
Частный скот |
0 |
Собственные данные |
Россия, Брянск |
Голштинская |
0 |
Хаертдинов Р.А., и др., 1997 |
Татарстан |