Потенціалкеровані натрієві канали.
Потенціалкеровані іонні канали реагують на зміни напруженості електричного поля в мембрані конформаційними змінами білків, що утворюють іонний канал.
Концепція їх існування зародилась ще на початку 50 – х років минулого століття з розшифровкою іонних механізмів генерації потенціалу дії. Це значною мірою зумовило той факт, що історично класичні потенціалзалежні іонні канали названо по типу основного іона який вони переносять – Na+, K+, Ca2+.
Головною особливістю потенціалзалежних натрієвих, калієвих та кальцієвих каналів є те, що їх функціональний стан визначається величиною мембранного потенціалу.
Будова та властивості потенціалкерованих натрієвих каналів;
Канал складається з α – субодиниці (Мм = 260 кДА), що являє собою трансмембранний білок. α – субодиниця складається із чотирьох доменів I – IV, кожен домен із шести сегментів, що формуються а – спіралями. До неї приєднано кілька вуглеводних ланцюгів, і вона має рецептор для зв'язування токсинів на зовнішній поверхні, а також локуси для фосфорилювання протеїнкіназами на внутрішньоклітинній поверхні. α – субодиниця асоційована з двома поліпептидами: β1 (Мм = 36 кДа), β2 (Мм = 33 кДа).
β1, β2 -субодиниці є глікозильованими. β1 - субодиниця нековалентно асоційована з а-субодиницею, тоді як β2 -субодиниця поєднана з нею дисульфідним зв'язком. Встановлено, що β1, β2 -субодиниці являють собою інтегральні мембранні глікопротеїни, тобто їхня гідрофобна частина розташована в подвійному ліпідному шарі. Значна кількість вуглеводів на субодиницях каналу припадає на сіалову кислоту, яка створює негативний поверхневий заряд.
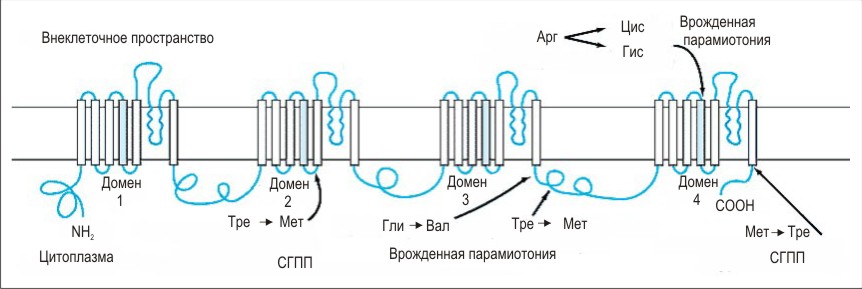
Рис. 7. Схема будови потенціал керованих натрієвих каналів.
Модулятори натрієвих каналів:
Існують два класи токсинів, які або спонтанно відкривають натрієві канали, або запобігають їх нормальному закриттю після того, як їх було відкрито. Це розчинні в ліпідах стероїди, такі як рослинні алкалоїди:
вератридин,
аконітин,
батрахотоксин - один з токсинів, та пептидні токсини отрути скорпіонів і анемон, які діють на рецептор на зовнішній поверхні мембрани.
Серед блокаторів натрієвих каналів особливе місце посідають
тетродотоксин
сакситоксин, які зв'язуються з рецепторним локусом, розташованим у зовнішньому вході натрієвого каналу, і блокують провідність унаслідок оклюзії пори.
Лідокаїн і прокаїн (новокаїн) є широко відомими місцевими анестетиками, що застосовують у клініці. Потенціалзалежними блокаторами натрієвих каналів є також низка фармакологічних препаратів, які застосовують у клініці як антиаритміки й протисудомні засоби.
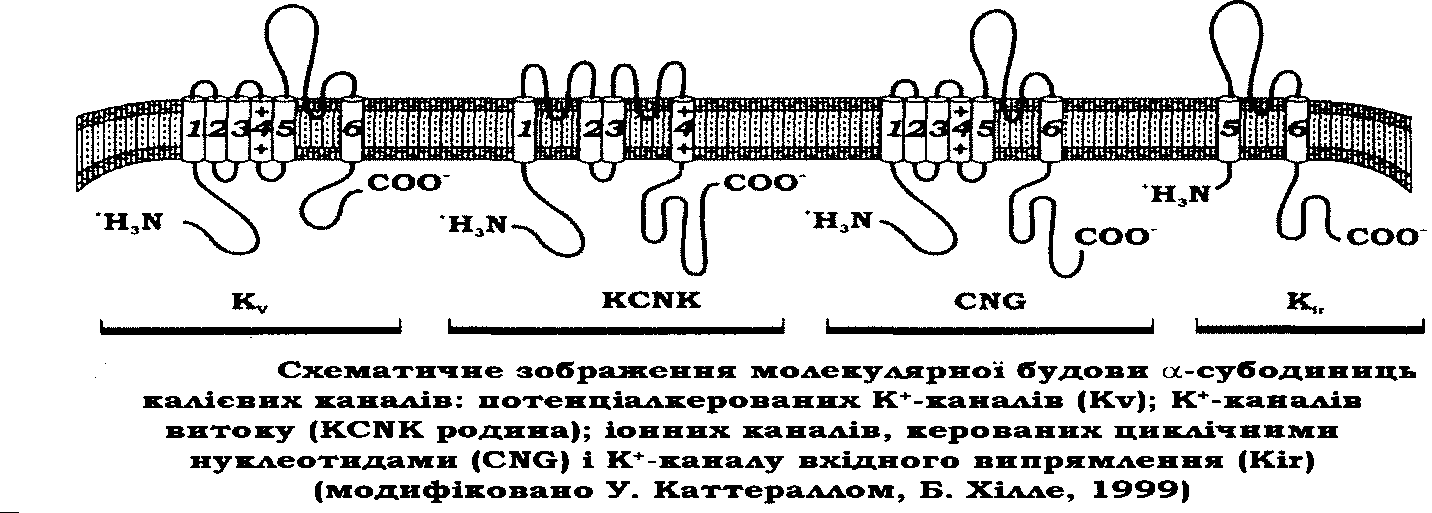
2. Калієві канали Загальна характеристика
К+ - канали - це велика група макромолекул, що формують пори, які переважно є проникними для К+.
Вони відіграють значну роль у різних функціях клітини. К+ - канали беруть участь у регуляції мембранного потенціалу і, таким чином, модулюють електричну збудливість клітин й електрохімічну рушійну силу для різноманітних транспортних механізмів. Роль потенціалкерованих К+ - каналів є важливою у формуванні змін мембранного потенціалу під час потенціалу дії в усіх збудливих клітинах. Вони функціонують у клітинних механізмах навчання та пам'яті.
У регуляції К+ - каналів беруть участь ковалентна модифікація, зумовлена фосфорилюванням, а також нековалентні взаємодії з іонами, нуклеотидами, киснем, ліпідами, такими зарядженими сполуками, як спермін, який належить до поліамінів.
За останні 10 років було клоновано десятки різновидів К+ - каналів.