

2.5. Генетико-прогностическое описание сс
2.5. Эволюция развития сенсоров у биологических организмов
3.1.1. Систематика
Стороннему наблюдателю может показаться удивительным, что биологическая систематика до сих пор не завершена и не согласована. Даже число крупнейших групп (типов), на которые подразделяется царство животных, остается предметом противоречий. Кроме того, с введением Хеннигом в 50-е годы XX века не-эволюционной кладистической схемы классификации, систематики оказались в центре горячей полемики. К счастью, нам нет необходимости вступать на это «минное поле». Наш интерес к положению организмов в классификации имеет выраженную эволюционную природу. Наш интерес к органам чувств животных, а не только нашим собственным, обусловлен не просто тем, что они существуют (хотя и это вполне уважительная причина), но главным образом тем, что их изучение способно пролить свет на сенсорные системы человека, а также продемонстрировать альтернативные пути решения тех же самых проблем. В некоторых случаях – это радикально иные решения (глаза насекомых и позвоночных), в других – поразительно схожие (глаза головоногих и позвоночных). Понимание эволюционных связей между группами животных – это основа полного понимания сенсорных систем животных и, в частности, тех, которые приобрели мы сами.
3.1.2. Классификация на шесть царств
Мы будем использовать общепринятую схему классификации организмов на шесть царств: Archaebacteria (архебактерии), Eubacteria (собственно бактерии), Protista (протисты), Plantae (растения), Fungi (грибы) и Animalia (животные). Внутри этих шести царств организмы подразделяются далее на большие группы со сходным планом строения, которые называются типами. Такой принцип классификации, как и можно было ожидать, куда более очевиден на многоклеточных – грибах, растениях и животных, чем в трех царствах одноклеточных. Действительно, классификация оказывается общепринятой только для царства животных. В изучении сенсорных систем именно оно занимает доминирующее положение, потому и наше внимание по большей части будет сконцентрировано на нем.
3.1.3. Одноклеточные
Archaebacteria и Eubacteria часто объединяются в единую группу «бактерий» или, иногда, монер (Moneres – греч., одинокий). Известно свыше 5 000 видов, представляющих собой маленькие клетки без ядра и органоидов. Поскольку эти клетки не имеют ясно выраженного ядра (т. е. нуклеиновые кислоты не отделены от цитоплазмы ядерной мембраной), они получили наименование прокариот. В течение прибл. 2 миллиардов лет (начиная с 3 600 млн лет назад до 1 600 млн лет назад) монеры владели миром. Сенсорные системы бактерий, конечно, предельно просты. Однако они представляют для нас значительный интерес, поскольку на них часто можно проследить, по крайней мере на биохимическом уровне, зачатки тех бесконечно сложных систем, которые развились у животных.
Спустя 2 млрд лет появились более крупные и более сложные клетки – эукариоты. Вероятно, они более тесно связаны с архебактериями, чем с собственно бактериями. Все эукариоты имеют выраженное ядро (их генетический материал сосредоточен за ядерной мембраной) и органоиды, такие как митохондрии, хлоропласты и т. д., локализованные в цитоплазме. Ранние классификации объединяли автотрофных эукариот с растениями в Prophyta, а гетеротрофных – с животными в Protozoa (простейшие). Более современный альтернативный анализ предполагает, что одноклеточные эукариоты схожи друг с другом значительно больше, чем с многоклеточными животными, растениями и грибами, и к тому же сама многоклеточность могла возникать несколько раз. Исходя из этого одноклеточные эукариоты и объединены в отдельное царство Protista или Protoctista (protos – первичный, kristos – устанавливать, греч.). Протисты – чрезвычайно разнообразная группа, подразделяющаяся на типы, которых насчитывается до 30, однако, в этой книге они, в основном, не фигурируют. С нашей точки зрения, самое интересное, возможно, заключается в наличии у них наиболее ранних фоторецепторов – различных типов светочувствительных пятен. Они обнаружены в типе Eustigmatophyta, Chlorophyta и, особенно, Euglenophyta (Euglena viridis), а также у некоторых представителей динофлагеллят (Pouchetia sp., Erythropsidinium pavillardii).

Рис. 4.1. (А) (а) Спиральное и (б) радиальное дробление. По Caroll, 1988. (Б) Первичноротые и вторичноротые. (а) Бластула; (б) инвагинация при формировании гаструлы. Бластоцель в основном смыкается, и образуется первичная кишка или архэнтерон, открытый во внешнюю среду (бластопор). У первичноротых он в дальнейшем формирует рот, а у вторичноротых возникает еще одна инвагинация (на противоположном от бластопора конце эмбриона), которая прорывается в архэнтерон. Здесь формируется рот, тогда как по соседству (рядом с зарастающим бластопором) прорывается вторичный анус.
3.1.4. Многоклеточность
Многоклеточность, вероятно, возникла около 1 млрд лет назад. Биологи выделяют три царства многоклеточных – грибы, растения и животные. Только последнее представляет интерес для данной работы. Здесь тоже имеется несколько различных методов классификации. В старых работах царство животных подразделялось на три полцарства: Protozoa (простейшие), Parazoa (Mesozoa, губки) и Metazoa (истинно многоклеточные). Позже, как мы видели, Protozoa были объединены с другими одноклеточными формами в царство Protista. Animalia (животные), в свою очередь, подразделяются на два под-царства – Parazoa и Metazoa. Parazoa далее подразделяются на Porifera (губки) и Placozoa (с единственным видом Trichoplax adhaerans, размером с большую амебу). Все остальные животные образуют Metazoa – многоклеточных. Старая (в высшей степени антропоцентрическая) классификация делит многоклеточных на позвоночных и беспозвоночных – в зависимости от наличия или отсутствия позвоночного столба. Такая классификация остается общепризнанной и общеупотребительной, но необъективной из-за нашего самомнения. Позвоночные (если быть более точным – черепные) образуют два подтипа внутри типа Chordata (хордовые), одного из более чем тридцати в подцарстве Metazoa.
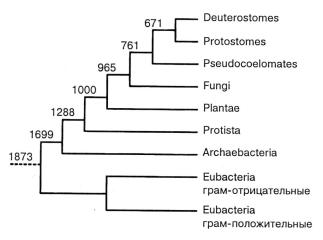
Рис. 4.2. Эволюционные взаимоотношения между основными группами организмов. Числами (в млн лет) показана давность дивергенций. Псевдоце-ломические животные представлены нематодами (напр. С. elegans).
3.1.5. Первичноротые и вторичноротые
Если внимательно рассмотреть ранний эмбриогенез целомических многоклеточных, то мы обнаружим несколько фундаментальных черт, которые предполагают разделение их на две большие группы форм животных: ветвь кольчатых червей – членистоногих – моллюсков (КЧМ) и ветвь иглокожих – хордовых (ИХ) (рис. 4.3). У кольчатых червей – членистоногих – моллюсков первые деления зигот (деления дробления) имеют спиральный характер (рис. 4.1 А(а)). Слои дочерних клеток (бластомеров) выстраиваются со сдвигом относительно друг друга. Проспективная судьба клеток таких зародышей детерминирована. Если одна из них удалена, соответствующая часть организма не сформируется. Напротив, в линии развития иглокожих – хордовых зигота претерпевает радиальное дробление (рис. 4.1 А(б)), а слои клеток расположены строго один над (под) другим. Судьбы бластомеров на этой стадии строго не детерминированы. Если один из них удален, остальные способны это компенсировать, так что формируется нормальный зародыш.
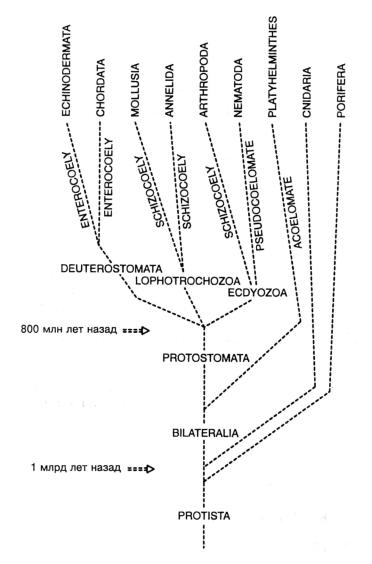
Рис. 4.3. Эволюционное древо царства животных. Всего насчитывается около 30 типов, но на рисунке показаны только те, которые включают виды, упоминающиеся на последующих страницах этой книги. Возникновение царства из протестов произошло около 1 млрд лет назад. Некоторые важные стадии формирования плана строения показаны на основном стволе схемы: билатеральная симметрия (BILATERALIA); выраженная полость тела (целом), отделяющая кишку от мускулатуры (COELO-МАТА); развитие рта из бластопора (PROSTOMA-ТА – первичноротые) или на противоположном конце (DEUTEROSTOMATA -вторичноротые). Все эти важные события эволюции имели место в докембрийский период.
Далее, две эти большие ветви демонстрируют глубокие различия путей формирования целома (полости тела). В линии КЧМ это происходит в результате расщепления мезодермы (шизоцель). В линии ИХ это происходит в результате отшнуровки выпячиваний кишки (энтероцель).
Наконец, важное различие – в том, как формируется рот. Сфера из клеток, образующаяся в результате делений дробления оплодотворенной яйцеклетки – бластула – затем инвагинирует, формируя первичную кишку – архэнтерон. Этот процесс называется гаструляцией. В ветви КЧМ отверстие первичной кишки – бластопор – в конце концов развивается в рот взрослой особи. Животные, у которых рот развивается таким образом, относятся к первичноротым. Напротив, в ветви ИХ рот развивается иначе. В этом случае новое отверстие в конце концов возникает на конце, противоположном бластопору, а бластопор преобразуется в анус (рис. 4.1 А(б)). Животные с таким типом развития относятся к вторичноротым (поскольку рот развивается у них «вторично»). Эти два термина – первичноротые и вторичноротые – обычно употребляются для обозначения двух упомянутых выше крупных ветвей развития.
Хотя т. н. «биогенетический закон» Геккеля (онтогенез отражает филогенез), в настоящее время принимается со значительными оговорками, нет сомнений, что последовательность событий раннего эмбриогенеза сложилась в эволюционной истории очень давно. Из этого ясно, что и принципиальные различия эмбриогенеза первично- и вторичноротых возникли в филогенезе очень рано. Молекулярно-биологические данные предполагают, что их общий предок существовал более, чем 670 млн лет назад. Недавние работы отодвинули этот срок на еще более давний срок – порядка 830 млн лет. Таким образом, эти две ветви в течение более трех четвертей миллиарда лет развиваются независимо друг от друга. Это делает возможность сравнения органов чувств между этими ветвями особенно интересной – нам нет необходимости лететь на другую планету, чтобы получить возможность сравнения с эволюционными альтернативами форм жизни, основанных на углероде.
В заключение, накапливаются молекулярно-биологические данные, дающие основания полагать, что первичноротые очень рано разделились на две крупных группы. Первая из них – Ecdyozoa – включающая, помимо прочих, нематод и членистоногих и характеризующаяся твердым наружным покровом, что требует периодических линек (ecdysis) для обеспечения возможности роста. Другая – Lophotrochozoa – жестким покровом не обладает (что, правда, отрицательно сказывается на их защищенности) и включает плоских и кольчатых червей, моллюсков, а также несколько других типов животных.