

8. 6. Экспрессия генов в бактериальных клетках и микроорганизмах
Основной единицей наследственности любого организма являются гены. Они представляют собой участки молекулы ДНК, расположенной в хромосоме. В простейшем случае - один ген содержит информацию о структуре одного белка, в более сложном – о нескольких белках. Информация о генах, их положении в геноме и методах получения важна для генетической инженерии.
Гены определяют все характерные признаки и свойства живых организмов. При половом размножении образуется рекомбинантная ДНК, которая содержит генетическую информацию обоих родителей. Генная инженерия позволяет создать организмы с новыми наследственными признаками. Для достижения этого необходимо встроить (клонировать) природный или искусственно синтезированный ген в вирусы, бактериальную плазмиду или любые другие векторные системы, а затем перенести в геном хозяина-реципиента таким образом, чтобы встроенный ген сохранял способность экспрессировать белок (рис. 8.12).
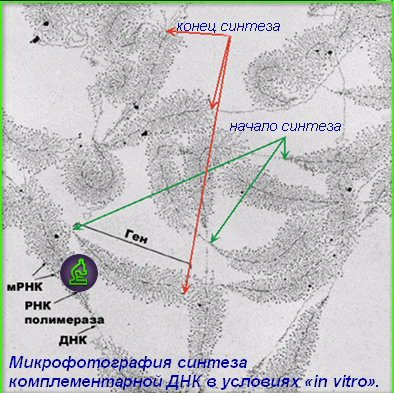
Рис. 8.12. Синтезирование гена для последующего встраивания его в векторные системы
Процесс разделения геномной ДНК на элементы и введение этих элементов в клетки-хозяева называется созданием геномной библиотеки (банки генов, банки клонов). Один из способов создания геномной библиотеки ДНК состоит в обработке донорской ДНК рестриктазой. Гидролиз проводят в мягких условиях, чтобы происходило лишь частичное расщепление. В результате образуются ферменты разных размеров, однако крупные не используются. Полная библиотека содержит весь геном данного организма.
При рестрикции ДНК образуется смесь разнообразных фрагментов, и после их лигирования с векторной ДНК возникает множество различных комбинаций. Необходимо уметь распознавать те реципиентные клетки, которые содержат ДНК с нужной нуклеотидной последовательностью. Для этого используют различные системы скрининга (поиски генов).
Следовательно, один ген содержит информацию о структуре одного белка, в более сложном варианте – о нескольких белках. Такая информация о положении генов в геноме важна для генетической инженерии.
Экспрессия гена – реализация генетической информации, закодированной в ДНК, путём её транскрипции и трансляции и-РНК.
Механизмы экспрессии генов у прокариот сегодня хорошо изучены и понятны благодаря многолетним фундаментальным молекулярно-биологическим исследованиям бактерии E. coli и некоторых её фагов. Начальным этапом экспрессии является процесс транскрипции, при котором фрагмент РНК-полимераза распознаёт на ДНК специфические последовательности, называемыми промоторами (участки генов, ответственных за начало их транскрипции). Присоединившись к ДНК в области промотора, РНК-полимераза расплетает двойную спираль ДНК и в присутствии субстратов начинает синтезировать нить м-РНК, комплементарную одной из нитей ДНК. Копируемая нить называется «значащей» нитью. При этом при одинаковых условиях с одних промоторов (сильных) синтез РНК начинается часто, с других (слабых) – редко.
В настоящее время расшифрованы нуклеотидные последовательности более 100 промоторов из E. coli и её фагов и многие десятки – из других бактерий. Установлено, что нуклеотидные последовательности в консервативных областях (районы «-10» и «-35»; рис. 8.13) и расстояние между ними определяют силу промотора.
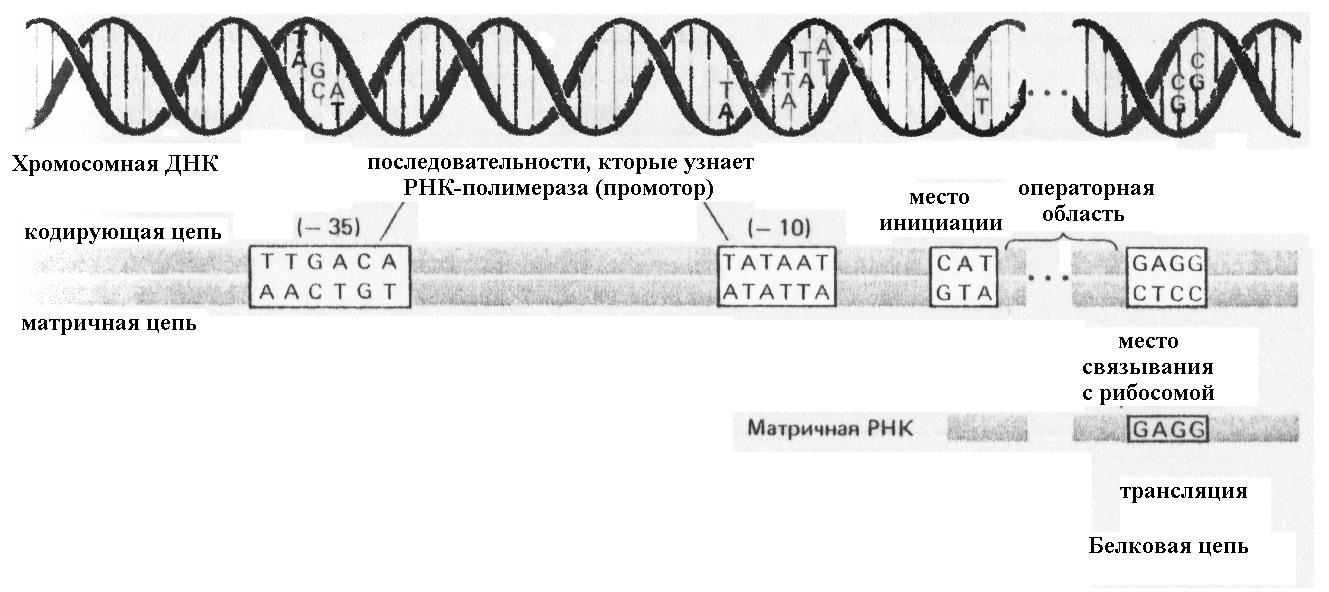
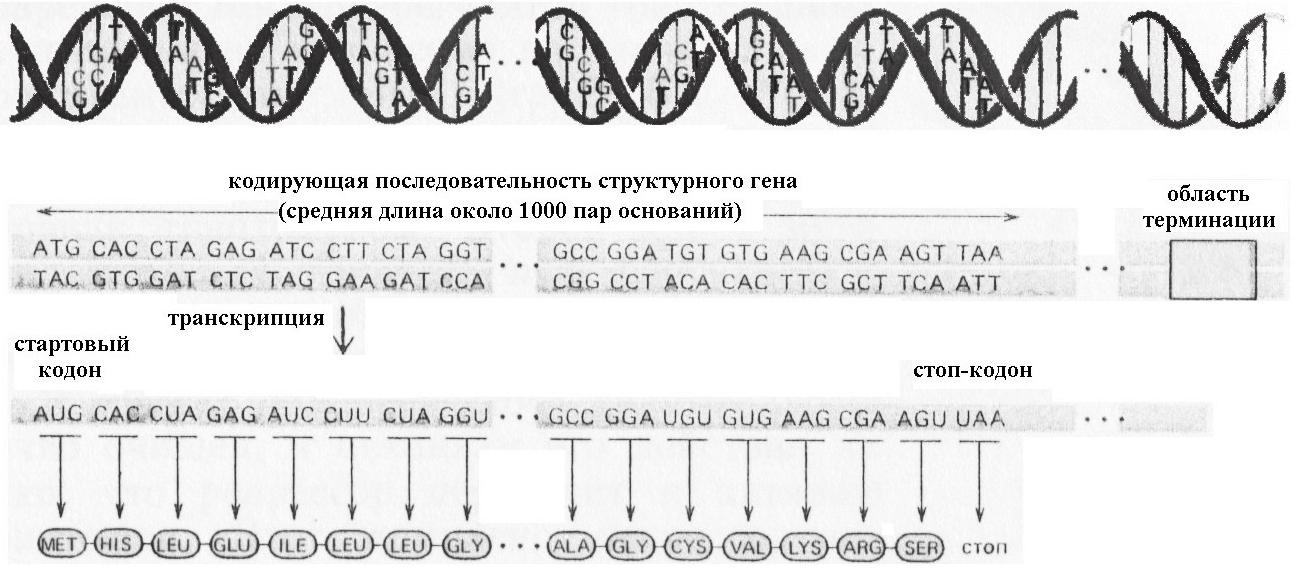
Рис. 8.13. Реализация генетической информации у бактерий
Видно, что даже в консервативных областях, например в области «- 10 » имеется усреднённая последовательность для E. coli – ТАТААТ, или в области «-35 » (усреднённая последовательность ТТGCA), у разных промоторов наблюдаются отличия по 1-3 нуклеотидам.
Промоторы разных бактерий близки по строению, что подтверждается не только структурой, но и их способностью инициировать транскрипцию при переносе в другой вид бактерий. При таком переносе эффективность работы промотора может быть снижена в десятки раз.
Поскольку эффективность определяется строением и специфичностью РНК-полимеразы клетки хозяина, то практически все гены бацилл, перенесённые в клетки E.coli вместе со своими промоторами, экспрессируются в этом организме. Вместе с тем, почти все гены E. coli, перенесённые в Bac. subtilis, не экспрессируются. Это означает более широкую специфичность РНК-полимеразы E.coli по сравнению с РНК-полимеразой Bac. subtilis.
Следовательно, первым условием экспрессии чужеродного гена в микроорганизмах должно быть наличие перед этим геном сильного промотора, распознаваемого РНК-полимеразой клетки-хозяина.
Для практических целей удобно использовать регулируемую экспрессию, так как не только сверхсинтез РНК, но и особенно сверхсинтез многих белков может оказаться гибельным для клетки. При таком подходе в процессе периодической ферментации в первой её фазе, когда происходит рост клеток и накопление биомассы, экспрессия клонированных чужеродных генов не происходит. Затем на втором этапе внешний индуктор (химическое вещество, температура) запускает сверхсинтез нужного белка.
Перенос генетической информации от чужеродной ДНК к белкам является основой экспрессии генов, которая осуществляется благодаря реализации их транскрипции и трансляции.
Транскрипция.
Транскрипция, или процесс синтеза РНК по матрице ДНК, определяется степенью сродства фермента РНК-полимеразы к промотору, представляющему последовательность нуклеотидных пар, опознаваемую РНК-полимеразой (рис. 8.14). Сам синтез белка на РНК, происходящий в рибосомах, зависит от стабильности РНК и её способности соединиться с рибосомами для образования белка. Такая последовательность передачи наследственной информации по пути: ДНК » РНК » белок – универсальна.
4 2 1 3 5
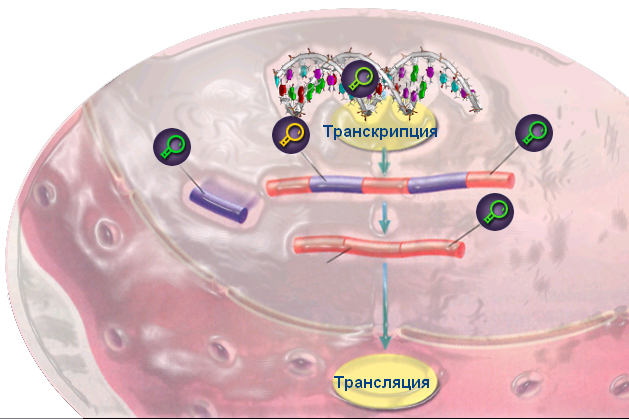
Рис. 8.14. Схема транскрипции РНК-полимеразы с промотором