

11. 3. Нейро-гуморальная регуляция внутрияичниковых процессов. Рост и развитие эмбрионов
Схема механизма нейро-гуморальной регуляции внутрияичниковых процессов у самок сельскохозяйственных животных представлен на рис. 11.3.
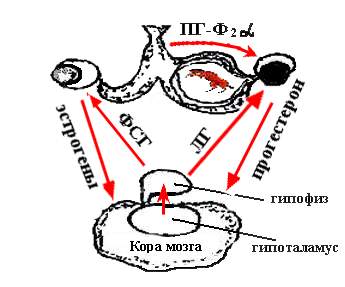
Рис. 11.3. Механизм нейро-гуморальной регуляции
Внутрияичниковых процессов
При отсутствии в матке зародыша, эндометрий выделяет в конце полового цикла простагландин Ф-2α, который, действуя на яичник, вызывает регрессию желтого тела. При этом размножившиеся клетки желтого тела перерождаются и рассасываются, превращаясь в небольшие бесцветные тельца, т.е. происходит фаза обратного развития. Лишь после этого наблюдается активный рост и созревание новых фолликулов.
При отсутствии в матке зародыша, эндометрий выделяет в конце полового цикла простагландин Ф-2α, который, действуя на яичник, вызывает регрессию желтого тела. При этом размножившиеся клетки желтого тела перерождаются и рассасываются, превращаясь в небольшие бесцветные тельца, т.е. происходит фаза обратного развития. Лишь после этого наблюдается активный рост и созревание новых фолликулов.
Эстрогены усиливают кровообращение и секрецию слизи в половых органах. В среднем мозгу возникает очаг возбуждения, что приводит к проявлению клинических признаков течки, полового возбуждения и охоты. В это время созревающие фолликулы продуцируют возрастающее количество эстрадиола, особенно в предовуляторный период. Это обуславливает повышение их чувствительности к гонадотропинам крови и обеспечивает дальнейшее развитие фолликулов до овуляции. Примерно, между 15 и 18-м днями полового цикла у самок отмечается резкое уменьшение содержания прогестерона в крови и увеличение эстрогенов.
Повышенное содержания эстрадиола активизирует функцию гипоталамуса, который, в свою очередь, стимулирует выделение из передней доли гипофиза «овуляторного» количества лютеинизирующего гормона (ЛГ). Стенка фолликула истончается и происходит ее разрыв под влиянием гидростатистического давления фолликулярной жидкости. Обычно овуляция происходит спустя 12,5 ч после окончания охоты. На месте лопнувшего фолликула образуется желтое тело, секретирующее прогестерон, которое ректальным методом хорошо пальпируется с 4-го дня после овуляции. Быстрое увеличение концентрации в крови прогестерона угнетает секрецию гонадотропин-рилизинг гормона (Гн-РГ) и препятствует дальнейшему освобождению ЛГ. Овуляция новых фолликулов не происходит.
Под влиянием прогестерона эндометрий полностью подготавливается к приему и прикреплению зиготы и обеспечивает нормальное течение беременности.
Рост и развитие эмбрионов в половых органах коровы-донора.
Локализация и стадия развития оплодотворенных яйцеклеток имеют особо важное значение, при проведении работы по извлечению эмбрионов из половых органов при трансплантации эмбрионов (рис. 11.4).
Началом оплодотворения яйцеклетки считается день, когда проведено первое осеменение коровы- донора. Этот день обозначается нулем (0), и начинается отсчет развития эмбриона до его извлечения.
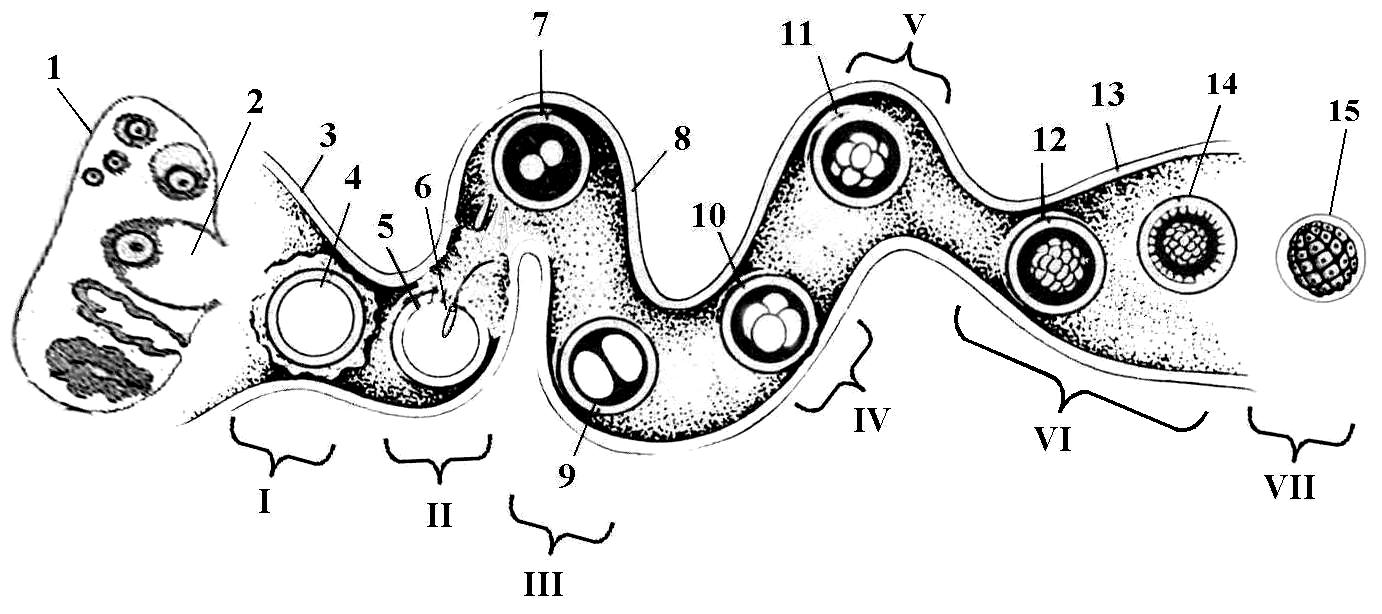
Рис.11.4. Стадии развития эмбрионов коров
I – нулевой день полового цикла; II – 1-й день после овуляции (оплодотворение); III – 2-й день после овуляции; IV – 3-й день после овуляции; V – 5-й день после овуляции; VI – 6-й день после овуляции; VII – 6-й день после овуляции.
1. Яичник; 2. Лопнувший зрелый фолликул; 3. Воронка яйцевода; 4. Яйцеклетка с лучистым венцом; 5. Яйцеклетка, освободившаяся от лучистого венца; 6. Проникновение спермия через прозрачную оболочку; 7. Образование зиготы (слияние женского и мужского ядер); 8. Яйцевод; 9. Стадия 2-х бластомеров; 10. Стадия 4-х бластомеров; 11. Стадия 8-ми бластомеров; 12. Стадия 16-ти бластомеров; 13. Верхушка рога матки; 14. Стадия 32-х бластомеров (ранняя морула); 15. Стадия 64-х бластомеров (поздняя морула).
До образования морулы ежедневно, через каждые 24 ч, происходит удвоение бластомеров. Так, на 2-ой день после овуляции образуется два бластомера, на 3-й - четыре, на 4-й – восемь и на 5-й день уже сформирован 16- клеточный эмбрион. При переходе в матку на, 4-й день, большинство эмбрионов находятся на 16-32-х клеточной стадии деления бластомеров. На 7 день (время извлечения эмбрионов) большинство достигает стадии поздней морулы (более 64) или ранней бластоцисты (130 бластомеров).
В это время они все находятся в концах рогов матки и, поэтому, их без труда можно извлечь вместе с промывной жидкостью с помощью катетера Фоллея.
Началом оплодотворения яйцеклетки считается день, когда проведено первое осеменение коровы- донора. Этот день обозначается нулем (0), и начинается отсчет развития эмбриона до его извлечения.
До образования морулы ежедневно, через каждые 24 ч, происходит удвоение бластомеров. Так, на 2-ой день после овуляции образуется два бластомера, на 3-й - четыре, на 4-й – восемь и на 5-й день уже сформирован 16- клеточный эмбрион. При переходе в матку на, 4-й день, большинство эмбрионов находятся на 16-32-х клеточной стадии деления бластомеров. На 7 день (время извлечения эмбрионов) большинство достигает стадии поздней морулы (более 64) или ранней бластоцисты (130 бластомеров).
В это время они все находятся в матке и их без труда можно извлечь вместе с промывной жидкостью с помощью катетера Фоллея.