

8.3. Ферменты в генной инженерии
Важная роль в проведении генно-инженерных работ принадлежит ферментам, участвующим в формировании рекомбинантных ДНК.
Цель создания рекомбинантной ДНК (т. е. введения чужого гена): наделить новый организм не имевшимися ранее наследственными свойствами. Для этой цели, прежде всего, следует сконструировать фрагмент чужеродной ДНК, содержащей этот столь необходимый ген.
Для этого, с помощью ферментов рестриктазы и лигазы «разрезают» и «сшивают» участки ДНК. Каждая рестриктаза, действуя на двухцепочную ДНК, «распознаёт» в ней только свою определённую последовательность нуклеотидов, прикрепляется к ней и разрезает её в месте прикрепления.
1. Рестриктазы одинаково эффективно разрезают ДНК человека, животного или растения, бактерии или вируса, лишь бы в ней были распознаваемые участки. В принципе, имеется в виду, что две совершенно несхожих между собой последовательности ДНК, например, из клеток мыши и кошки, при обработке одной и той же рестриктазой можно эффективно сшить друг с другом.
Традиционно, рестриктазы распознают в молекулах ДНК очень короткие, но строго специфичные для каждого фермента участки длиной в 4-6 пар нуклеотидов и разрезают обе цепи ДНК: 1. Посередине этих участков; 2. С некоторым смещением. В первом случае образуются обрывки с ровными (тупыми) концами, а во втором – стороны разрезаемых цепочек ДНК заходят одна за другую (рис. 8.6). Такие одноцепочные концы называются «липкими», поскольку они могут слипаться между собой в силу комплементарности.
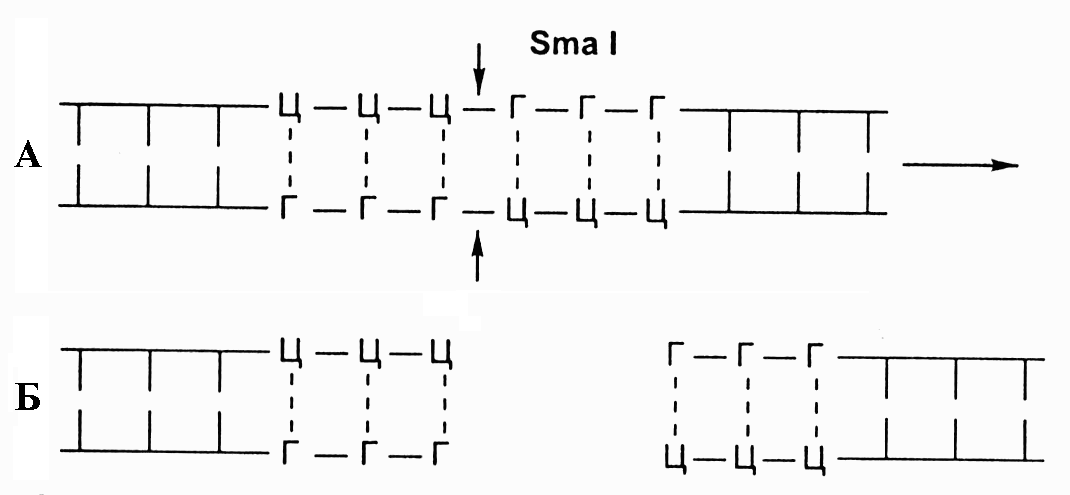
Рис. 8.6. А - схема действия фермента рестриктазы Sma I
на двухцепочечную молекулу ДНК, с указанием участка
распознавания и места разреза; Б - фрагменты ДНК с тупыми концами после разрезания ферментом Sma I
Примером действия рестриктазы второго приведённого случая является EcoRI, которая узнаёт фрагмент ДНК из шести нуклеотидов ГААТТЦ и режет эту последовательность ДНК ассиметрично, «ступенькой», между нуклеотидами Г и А. (рис. 8. 7).
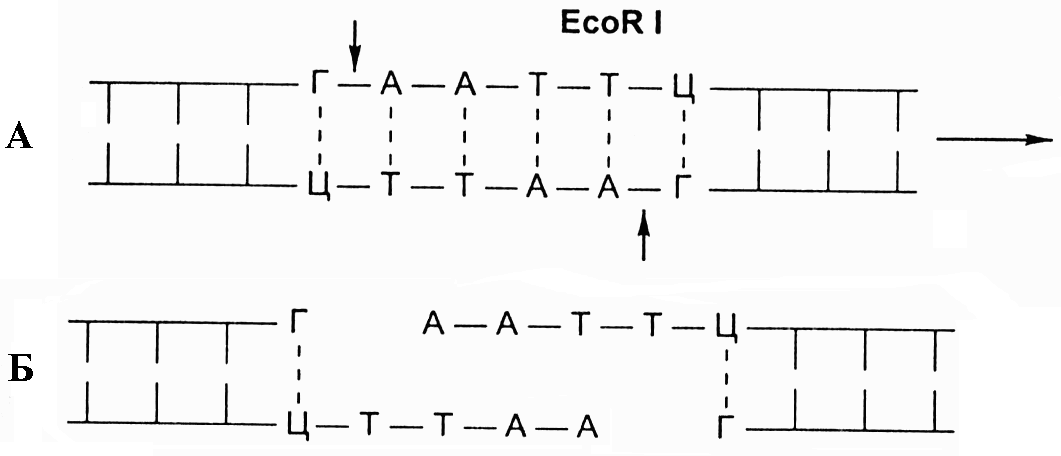
Рис. 8.7. А - схема действия фермента рестриктазы EcoRI
на двухцепочечную молекулу ДНК, с указанием участка распознавания и места разреза; Б - фрагменты ДНК
с липкими концами после разрезания ферментом EcoRI
В результате место разреза в одной цепи смещено по отношению к другой на 4 пары оснований. При таком разрезе образуются два выступающих конца. Эти концы притягиваются друг к другу, желая восстановить свои старые связи и скрепиться, как им и положено, водородными мостиками. Если с той же EcoRI получить фрагменты ДНК из различных организмов, то все они будут иметь одинаковые, подходящие друг к другу «липкие концы». Он лигирует, т.е. «сшивает» между собой сахарофосфатные основы двух фрагментов с образованием полной структуры двойной спирали ДНК.
В настоящее время в практике генной инженерии применяется около 500 различных рестриктаз, способных разрезать ДНК примерно в 120 различных местах. Примеры рестриктаз и участков ДНК, которые они могут разрезать, представлены в табл. 8.3).
Таблица 8.3. Рестриктазы и расщепляемые ими последовательности

Данный пример показывает, как можно с помощью данных ферментов в условиях генно-инженерных лабораторий конструировать разнообразные по своим составным частям гибридные ДНК.
Таким образом, рестриктазы (рестрикционные нуклеазы) – это ферменты, способные узнавать специфические последовательности (4-6 нуклеотидов) ДНК и разрезать их в строго определенных местах. Когда два разных образца обрабатывают одной и той же рестриктазой, а затем смешивают эти образцы, то благодаря комплементарному спариванию фрагментов разных образцов могут образовываться новые комбинации генов – рекомбинантные ДНК.
2. ДНК-лигазы осуществляют «сшивание» (связывание) фрагментов ДНК, образуемых в процессе репликации за счет образования фосфодиэфирных связей между соседними нуклеотидами. Бактериальная ДНК-лигаза склеивает разрывы за счет «липких концов». Лигаза фага Т4 соединяет двунитевые фрагменты ДНК, содержащие ровные концы. Лигазы являются второй по важности в генетической инженерии группой ферментов.
Ниже представлен процесс сшивания фрагментов ДНК, содержащих нужные гены, с использованием двух основных методов:
1. По «липким» концам;
2. С помощью искусственно достроенных «липких» концов.
Сшивание генов (фрагментов) ДНК по «липким» концам комплементарных участков, длиной от 4 до 6 пар нуклеотидов, достаточно легко осуществляется ферментом ДНК-лигазой с образованием ковалентной связи между соседними нуклеотидами:
– – А Т Г Ц А А Т Т Ц А Г Т Ц – – – – – –
Т А Ц Г Т Т А А Г Т Ц А Г – – – – – –
Сшивание ДНК - лигаза

А Т Г Ц А А Т Т – Ц А Г Т Ц
Т А Ц Г– Т Т А А – Г Т Ц А Г
При отсутствии таких «липких» концов у сшиваемых фрагментов их достраивают, т. е. синтезируют искусственно ферментативным путём. Для этой цели применяют линкеры (или «переходники») – короткие участки ДНК, имеющие разные «липкие» концы:
– А Т Г Ц А А Т Т Ц Т Г А Г А Т Ц Ц А Т А Ц Г
– Т А Ц Г Т Т А А Г А Ц Т Ц Т А Г Г Т А Т Г Ц
Фрагмент 1 Линкер Фрагмент 2
Переходниковые линкерные фрагменты обеспечивают как объединение генов, так их экспрессию, для чего в середину линкера помещают какой-либо регуляторный генетический элемент, например промотор, или участок связывания с рибосомой.
Возможно сшивание фрагментов и по тупым концам, когда концы фрагментов двунитевые:
– – – А Т Г Ц Г Г Г Ц Ц Ц Г Т А – – – – – –
– – – Т А Ц Г Ц Ц Ц Г Г Г Ц А Т – – – – – –

В данном случае реакция лигирования (сшивания) имеет биохимические особенности и её эффективность ниже, чем при сшивании по «липким» концам.
Сшитая рекомбинантная ДНК вводится в живые клетки. Однако она не способна к самовоспроизведению, в связи с чем, её разрушают внутриклеточные нуклеазы. Для того чтобы рекомбинантная ДНК стала составной частью генетического аппарата клетки она должна:
1. Встроиться (интегрироваться) в геном клетки и реплицироваться, либо быть способной к автономной репликации молекулы ДНК;
2. Иметь способность автономно реплицироваться (называются векторными молекулами). К их числу относят плазмиды, бактериофаги, вирусы животных.
Используя ферменты рестриктаз и лигаз, на сегодняшний день решена проблема конструирования разнообразных по своим составным частям гибридных (рекомбинантных) ДНК, путём сшивания фрагментов разных видов in vitro. Основное назначение полученных гибридных генов попавших в клетку заключается в том, чтобы производить новые белки.
Рестрикционные карты.
На основании изучения фрагментов ДНК создают «Рестрикционные карты». На этих картах представлена последовательность всех нуклеотидов, входящих в состав ДНК, а также отмечены сайты (места) действия рестриктаз. Первая такая карта была получена для вируса SW40 (обезьяний вирус, вызывающий злокачественную трансформацию), содержащего 5423 пары оснований (рис. 8. 8).
Помимо рестриктаз и ДНК-лигаз в генно-инженерных работах используются и другие ферменты:
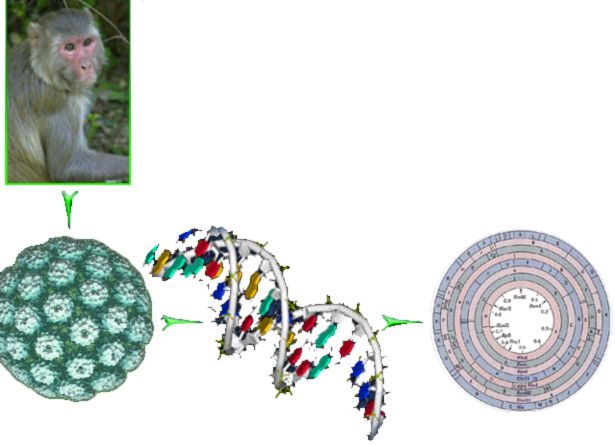
Рис. 8. 8. Первая рестрикционная карта получена для вируса SW40 (обезьяний вирус), содержащего 5423 пары оснований
3. Ревертаза. Ее функция связана с синтезом на матрице м-РНК.
4. ДНК и РНК-полимеразы обладают общим свойством вести матричный синтез нуклеиновых кислот в направлении 51 → 31.
5. Терминальная трансфераза – способна наращивать на концах фрагментов ДНК однонитевые участки путем последовательного присоединения нуклеотидных остатков из тех дезоксинуклеозидтрифосфатов, которые добавляются в реакционную смесь. Ее используют для создания на соединяемых фрагментах ДНК искусственных липких концов.
6. Нуклеазы – это большой класс ферментов, расщепляющих молекулы нуклеиновых кислот. Имеются нуклеазы, расщепляющие одно - или двуцепочные ДНК или РНК путем отщепления по одному нуклеотиду или небольших олигонуклеотидов.