

Гладкие мышцы
Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и сосудов является миоцит. Представляет собой чаще всего веретенообразную клетку (длиной 20-500 мкм, диаметром 5-8 мкм), покрытую снаружи базальной пластинкой, но встречаются и отростчатые миоциты. В центре располагается вытянутое ядро(Рис 15), по полюсам которого локализуются общие органеллы: зернистая эндоплазматическая сеть, пластинчатый комплекс, митохондрии, цитоцентр. В цитоплазме содержатся толстые (17 нм) миозиновые и тонкие (7 нм) актиновые миофиламенты, которые располагаются в основном параллельно друг другу вдоль оси миоцита и не образуют А и I диски, чем и объясняется отсутствие поперечной исчерченности миоцитов. В цитоплазме миоцитов и на внутренней поверхности плазмолеммы встречаются многочисленные плотные тельца, к которым прикрепляются актиновые, миозиновые, а так же промежуточные филаменты. Плазмолемма образует небольшие углубления - кавеолы, которые рассматриваются как аналоги Т-канальцев. Под плазмолеммой локализуются многочисленные везикулы, которые вместе с тонкими канальцами цитоплазмы являются элементами саркоплазматической сети.
Механизм сокращения в миоцитах в принципе сходен с сокращением саркомеров в миофибриллах в скелетных мышечных волокнах. Он осуществляется за счет взаимодействия и скольжения актиновых миофиламентов вдоль миозиновых. Для такого взаимодействия также необходимы энергия в виде АТФ, ионы кальция и наличие биопотенциала. Биопотенциалы поступают от эфферентных окончаний вегетативных нервных волокон непосредственно на миоциты или опосредованно от соседних клеток через щелевидные контакты и передаются через кавеолы на элементы саркоплазматической сети, обуславливая выход из них ионов кальция в саркоплазму. Под влиянием ионов кальция развиваются механизмы взаимодействия между актиновыми и миозиновыми филаментами, аналогичные тем, которые происходят в саркомерах скелетных мышечных волокон, в результате чего происходит скольжение названных миофиламентов и перемещение плотных телец в цитоплазме. В миоцитах, кроме актиновых и миозиновых филаментов, имеются еще промежуточные, которые одним концом прикрепляются к цитоплазматическим плотным тельцам, а другим - прикрепительным тельцам на плазмолемме и таким образом передают усилия взаимодействия актиновых и миозиновых филаментов на сарколемму миоцита, чем и достигается его укорочение.
Миоциты окружены снаружи рыхлой волокнистой соединительной тканью - эндомизием и связаны друг с другом боковыми поверхностями. При этом, в области тесного контакта соседних миоцитов базальные пластинки прерываются. Миоциты соприкасаются непосредственно плазмолеммами и в этих местах имеются щелевидные контакты, через которые осуществляется ионная связь и передача биопотенциала с одного миоцита на другой, что приводит к одновременному и содружественному их сокращению. Цепь миоцитов, объединенных механической и метаболической связью, составляет функциональное мышечное волокно. В эндомизии проходят кровеносные капилляры, обеспечивающие трофику миоцитов, а в прослойках соединительной ткани между пучками и слоями миоцитов в перимизии проходят более крупные сосуды и нервы, а также сосудистые и нервные сплетения.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой. При этом, терминальные веточки аксонов эфферентных вегетативных нейронов, проходя по поверхности нескольких миоцитов, образуют на них небольшие варикозные утолщения, которые несколько прогибают плазмолемму и образуют мионевральные синапсы. При поступлении нервных импульсов в синаптическую щель выделяются медиаторы (ацетилхолин или норадреналин), и обуславливают деполяризацию мембран миоцитов и последующее их сокращение. Через щелевидные контакты биопотенциалы переходят из одного миоцита на другой, что сопровождается возбуждением и сокращением и тех гладкомышечных клеток, которые не содержат нервных окончаний. Возбуждение и сокращение миоцитов обычно продолжительны и обеспечивают тоническое сокращение гладкой мышечной ткани сосудов и полых внутренних органов, в том числе гладкомышечных сфинктеров. В этих органах содержатся и многочисленные рецепторные окончания в виде кустиков, деревцев или диффузных полей.


1 2
Рис. 15. Гладкомышечные клетки:
1 – общий вид;
2 – А – в расслабленном состоянии; Б – в сокращенном состоянии; 1 – ядро; 2 – плотные тельца, прикрепленные к цитолемме (тельца прикрепления);
3 – промежуточные миофиламенты.
Автоматия. Многим внутренним органам и сосудам присуща автоматия гладких мышц, т.е. способность к автоматической (спонтанной) деятельности. Потенциал действия гладких мышечных клеток имеет авторитмический (пейсмекерный) характер, подобно потенциалам проводящей системы сердца. Пейсмекерные потенциалы регистрируются в различных участках гладкой мышцы. Это свидетельствует о том, что любые клетки висцеральных гладких мышц способны к самопроизвольной автоматической активности.
Реакция на растяжение. Уникальной особенностью клеток гладких мышц является их связь мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой.
В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
На рис. 16 представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки, где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце.
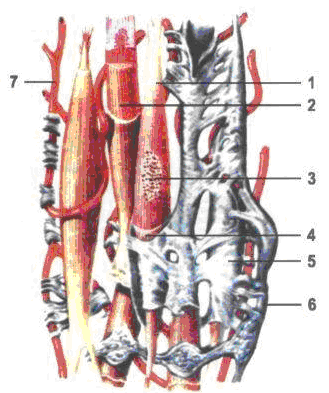
Рис. 16. Построение гладкой мышечной ткани: 1 – миоцит; 2 – миофибриллы в саркоплазме; 3 – ядро миоцита; 4 – сарколема; 5 – ендомизий; 6 – нерв; 7 – кровеносный капилляр.
Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой — в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины, в отличие от 30% укорочения, характерного для скелетной мышцы.
Висцеральные гладкие мышцы характеризуются нестабильным мембранным потенциалом. Колебания мембранного потенциала независимо от нервных влияний вызывают нерегулярные сокращения, которые поддерживают мышцу в состоянии постоянного частичного сокращения — тонуса. Тонус гладких мышц отчетливо выражен в сфинктерах полых органов: желчном и мочевом пузырях, в месте перехода желудка в двенадцатиперстную кишку и тонкой кишки в толстую, а также в гладких мышцах мелких артерий и артериол. Мембранный потенциал гладкомышечных клеток не является отражением истинной величины потенциала покоя. При уменьшении мембранного потенциала мышца сокращается, при увеличении — расслабляется. Платообразные потенциалы действия (ПД) обеспечивают поступление в цитоплазму миоцитов значительного количества внеклеточного кальция, участвующего в последующем в активации сократительных белков гладкомышечных клеток. Ионная природа ПД гладкой мышцы определяется особенностями каналов мембраны гладкой мышечной клетки. Основную роль в механизме возникновения ПД играют ионы Са2+. Кальциевые каналы мембраны гладких мышечных клеток пропускают не только ионы Са2+, но и другие двухзарядные ионы (Bа 2+, Mg2+), а также Na+. Вход Са2+ в клетку во время формирования ПД необходим для поддержания тонуса и развития сокращения. Поэтому блокирование кальциевых каналов мембраны гладких мышц, приводящее к ограничению висцеральной гладкой мышцы, является ее реакция на растяжение. В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение уменьшает мембранный потенциал клеток, увеличивает частоту ПД и в конечном итоге — тонус гладкой мускулатуры.
В организме человека это свойство гладкой мускулатуры служит одним из способов регуляции двигательной деятельности внутренних органов. Например, при наполнении желудка происходит растяжение его стенки. В ответ на его растяжение увеличивается тонус стенки желудка. Этоспособствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления, является основным фактором миогенной саморегуляции тонуса сосудов. Наконец, растяжение мускулатуры матки растущим плодом служит одной из причин начала родовой деятельности.
Пластичность. Еще одной важной специфической характеристикой гладкой мышцы является изменчивость напряжения без закономерной связи с ее длиной. Так, если растянуть висцеральную гладкую мышцу, то ее напряжение будет увеличиваться. Однако если мышцу удерживать в состоянии удлинения, вызванным растяжением, то напряжение будет постепенно уменьшаться. Иногда напряжение снижается не только до уровня, существовавшего до растяжения, но и ниже этого уровня. Это свойство называется пластичностью гладкой мышцы. Таким образом, гладкая мышцы более похожа на тягучую пластичную массу, чем на малоподатливую структурированную ткань. Пластичность гладкой мускулатуры способствует нормальному функционированию внутренних полых органов.
В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы. Эта особенность заключается в том, что прежде чем миозин гладкой мышцы сможет проявлять свою АТФазную активность, он должен быть фосфорилирован. Фосфорилирование и дефосфорилирование миозина наблюдается и в скелетной мышце, но в ней процесс фосфорилирования не является обязательным для активации АТФазной активности миозина.
Механизм фосфорилирования миозина гладкой мышцы осуществляется следующим образом: ион Са2+ соединяется с кальмодулином. Кальмодулин — рецептивный белок для иона Са2+. Возникающий комплекс активирует в легкой цепи миозина фермент — киназу, который в свою очередь катализирует процесс фосфорилирования миозина. Образуется актомиозиновый комплекс. Затем происходит скольжение актина по отношению к миозину, составляющее основу сокращения. Отметим, что триггером сокращения гладкой мышцы является присоединение иона Са2+ к кальмодулину. В скелетной же и сердечной мышце триггер – присоединение Са2+ к тропонину.
Химическая чувствительность. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину, ацетилхолину, гистамину и др. Это обусловлено наличием специфических рецепторов мембраны гладкомышечных клеток. Если добавить адреналин или норадреналин к препарату гладкой мышцы кишечника, то увеличивается мембранный потенциал, уменьшается частота ПД и мышца расслабляется, т. е. наблюдается тот же эффект, что и при возбуждении симпатических нервов.
Норадреналин действует на α- и β-адренорецепторы мембраны гладкомышечных клеток. Взаимодействие норадреналина с β-рецепторами уменьшает тонус мышцы в результате активации аденилатциклазы. Это приводит к образованию циклического АМФ и последующему увеличению связывания внутриклеточного Са2+. Воздействие норадреналина на α-рецепторы тормозит сокращение за счет увеличения выхода ионов Са2+ из мышечных клеток.
Ацетилхолин оказывает на мембранный потенциал и сокращение гладкой мускулатуры действие, противоположное действию норадреналина. Добавление ацетилхолина к препарату гладкой мышцы кишечника уменьшает мембранный потенциал и увеличивает частоту спонтанных ПД. В результате увеличивается тонус и возрастает частота ритмических сокращений, т. е. наблюдается тот же эффект, что и при возбуждении парасимпатических нервов. Ацетилхолин деполяризует мембрану, увеличивает ее проницаемость для Na+ и Са2+.