

§9.4. Нуклеационный механизм сворачивания
В 1973 г. Птицыным О.Б. была предложена гипотеза стадийного сворачивания белка (framework model – каркасная модель).
Согласно этой модели (рис. 9–8) на первых этапах сворачивания из развернутой цепи (1) образуются зародышевые α-спирали и β-шпильки.
Затем, на второй стадии, также быстро, происходит ассоциация элементов вторичной структуры, приводящая к образованию различных сочетаний: нескольких α-спиралей, нескольких β-тяжей и смешанных α–β ассоциатов (2).
На третьей стадии образуются специфические контакты между удаленными друг от друга участками аминокислотных последовательностей, пространственно сближенных в третичной структуре (3). Чтобы возникла уникальная пространственная структура белка, для каждого вида белка необходимо образование оптимального числа таких контактов. Если возникают случайные ошибочные контакты, то белок перестаивается до тех пор, пока ни образуется оптимальная структура контактов для данного белка. Формируется расплавленная глобула (3), отличающаяся от нативной глобулы меньшей упорядоченностью структуры, меньшей плотностью упаковки гидрофобного ядра молекулы. Расплавленная глобула имеет структуру близкую к структуре нативного белка, но еще не обладает необходимой функциональной активностью. Расплавленная глобула лабильна и легко слипается с другими расплавленными глобулами с образованием агрегатов, что уменьшает число молекул, находящихся на правильном пути сворачивания. Образование расплавленной глобулы происходит значительно быстрее, чем ее переход в форму нативной глобулы, так как этот переход связан с большим числом взаимосвязанных структурных перестроек, происходящих достаточно медленно. Кинетический интермедиат сворачивания белка (2) называется, соответственно, пред-расплавленной глобулой.
Последняя стадия (3→4) – «доводка» структуры белка, образование плотно упакованной активной глобулы – является самой длительной стадией сворачивания белка (от секунд до часов в зависимости от белка).
Поскольку вероятность агрегации белковых молекул сильно возрастает с повышением температуры, то для эффективного сворачивания при функциональных температурах необходимо использовать разбавленные растворы.
Пред-расплавленная и расплавленная глобулы являются промежуточными метастабильными состояниями белковой молекулы на пути самоорганизации in vitro от клубка до и нативного белка. Как уже говорилось, расплавленная глобула образуется быстро за несколько миллисекунд, а основное время уходит на образование плотноупакованной структуры.
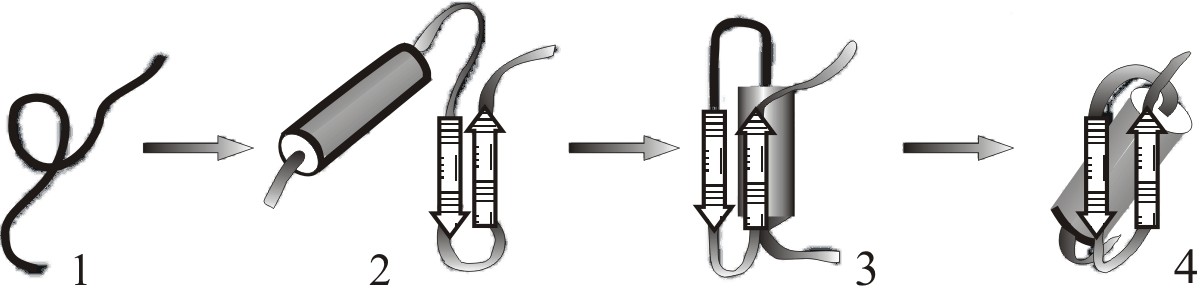
Рис. 9–8. Стадийная модель сворачивания по Птицыну: (1) - развернутая цепь; (2) - вторичные структуры – α-спирали (цилиндры) и β-листы (стрелки), флуктуирующие вблизи нативного положения; (3) – расплавленная глобула, обладающая вторичной структурой и мотивом укладки, подобным нативной структуре; (4) – нативная пространственная структура
Если сворачивание полипептидной цепи кодируется несколькими генами, то участки, кодируемые разными генами, сворачиваются независимо друг от друга, по разным путям и с разной скоростью. Образующиеся после сворачивания структуры называются доменами.
Экспериментов пока немного, но они подтверждают факт нуклеационного механизма сворачивания. При сворачивании белка определенные аминокислотные остатки группируются в компактный «доменчик», компактное ядрышко – зародыш сворачивания, причем оно лежит не в геометрическом центре белка, то есть не совпадает с гидрофобным ядром водорастворимых глобул, а сдвинуто к поверхности глобулы. Нуклеация типична для фазовых переходов первого рода. На пути сворачивания глобулы зародыш белковой структуры не перестраивается, все перестройки происходят только в рыхлом клубке и поэтому – быстро.
Большие белки (более 100÷150 остатков) состоят из отдельно сворачивающихся стабильных доменов («фолдонов», от fold - укладка). В многодоменных белках единицей самоорганизации является не белок в целом, а его отдельный домен.
Расчет показывает, что ядро сворачивания белковой глобулы достаточно большое. Оно охватывает около половины белковой глобулы. Столь большой размер ядра показывает, что белок не может сворачиваться по многим параллельным путям – путям с разными переходными состояниями и ядрами.