

Старение
Как только потомки половых или стволовых клеток начинают дифференцироваться, активность теломеразы падает, и их теломеры начинают укорачиваться. В полностью созревших дифференцированных клетках активность теломеразы падает до нуля, и с каждым делением клетки неотвратимо приближаются к состоянию сенесенса (одряхления, прекращения деления).
На рис. 9–29 приведены зависимости длины теломер (числа пар теломерных нуклеотидов) от числа делений клеток для зародышевых (эмбриональных), нормальных и раковых клеток. У зародышевых клеток длина теломер не изменяется при делении. У нормальных клеток длина уменьшается и стремится к лимиту Хейфлика. Затем клетки или гибнут, или превращаются в злокачественные (раковые). При переходе клеток в злокачественные длина теломер увеличивается настолько, чтобы обеспечить бесконечное деление клеток – иммортабельность (бессмертие). Сравнительно небольшая длина теломер у раковых клеток свидетельствует как раз о том, что они происходят из нормальных клеток, достигших предкризисного состояния.
Предкризисное состояние наступает в результате нарушения многих биохимических реакций, ведущих к многочисленным хромосомным перестройкам, в том числе, и к злокачественной трансформации. В настоящее время установлено, что для перерождения нормальной клетки в раковую в ней должно произойти (накопиться во времени) около 10 мутаций. Именно в результате случайных мутаций в клетке может активироваться теломераза, которая поддерживает длину теломер на уровне необходимом для их функционирования.
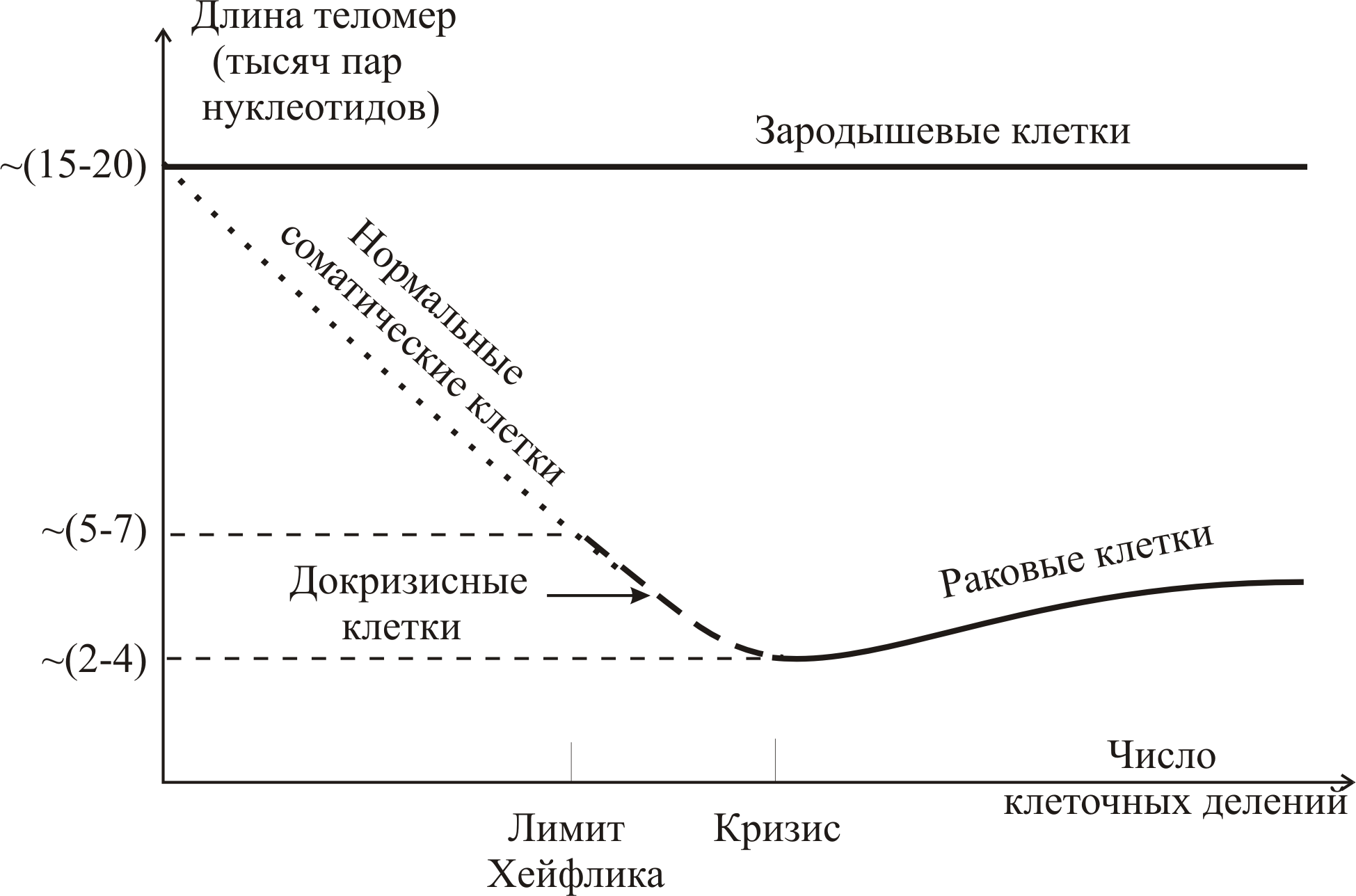
Рис. 9–29. Зависимость длины теломерной ДНК (в тысячах пар нуклеотидов) от числа клеточных делений. В зародышевых и раковых клетках теломераза активна (сплошные кривые), в нормальных соматических клетках теломеразная активность отсутствует (пунктирная и штриховая кривые), что приводит к уменьшению длины теломер с ростом числа делений клетки вплоть до критического значения. После чего клетка или погибает, или перерождается в раковую
Таким образом, избирательное подавление теломеразной активности в раковых клетках – это один из возможных способов борьбы со злокачественными заболеваниями.
О возможности продления жизни
В 1998 году было установлено, и это вызывало сенсацию, что нормальные соматические клетки потому лишены теломеразной активности, что в них полностью подавлена экспрессия (степень проявления гена, от лат. expressio – выражение) гена каталитической субъединицы теломеразы — обратной транскриптазы. Другие же составляющие теломеразы, включая теломеразную РНК, образуются в нужных количествах.
Отсюда следует, что если внести в нормальные клетки гены теломеразы (теломеразной обратной транскриптазы), то нормальные клетки должны стать "бессмертными". Этот вывод был подтвержден экспериментально. При введении в клетки указанных генов теломераза приобретала активность и способность поддерживать длину теломер на необходимом уровне. Клетки продолжали делиться до 220 раз и более, тогда как контрольная группа обычных "смертных" клеток делилась только 5060 раз.
9.8.4. Рак – генетическая болезнь