

Теломераза. Синтез теломеров
В отличие от рассмотренных выше дифференцированных клеток, зародышевые, стволовые и раковые клетки могут делиться неограниченное число раз. Это означает, что теломеры этих клеток сохраняют постоянную длину.
В 1984 г. Э. Блэкберн и Э. Грайдер выделили специальный фермент, синтезирующий теломерную ДНК. Этот фермент получил название теломеразы.
Основное назначение теломеразы — синтезировать повторяющиеся сегменты ДНК, из которых состоит цепь теломерной ДНК.
РНК, используемая теломеразой в качестве матрицы для синтеза теломерной ДНК, входит в состав фермента. Длина теломеразной РНК человека ~450 пар нуклеотидов. Матричный участок представлен в теломеразной РНК только один раз. Его длина не превышает длину двух повторов в теломерной ДНК, которую он кодирует. В качестве праймера при синтезе Г-цепи (верхней на рис. 9–28) используется участок ГАТ на 3´-конце хромосомной ДНК, с которым часть матричного участка теломеразной РНК образует комплементарный комплекс. Так как теломераза синтезирует сегменты ДНК, повторяющиеся много раз, используя только один сегмент (с 3’ конца: АУЦЦЦА) своей РНК, то она периодически (после завершения синтеза каждого повтора) перемещает матричный участок в район 3–конца Г-цепи синтезируемой теломерной ДНК (рис. 9–28), используя для этого энергию, выделяющуюся в процессе синтеза.
После завершения синтеза некоторый конечный участок (ТТАГ) на 3´-конце Г-цепи остается в однотяжном состоянии, чтобы обеспечить возможность наращивания на нем теломеразой нового повтора. Таким образом, механизм синтеза теломерной ДНК качественно отличается от репликации ДНК. Он осуществляется в два этапа. При каждом делении сначала теломераза достраивает однотяжевую теломерную цепь с 3´-конца до первоначальной длины, а затем ДНК-полимераза синтезирует на ней комплементарную Ц-цепь. Ц-цепь (5´-конец нижней цепи ДНК на рис. 9-28) синтезируется при участии обычной ДНК-полимеразы.
Высокая теломеразная активность наблюдается в половых клетках человека в течение всей его жизни. Соответственно их теломеры состоят из наибольшего числа повторов и содержат все необходимые белки для нормального деления клеток. Аналогичная ситуация наблюдается и для стволовых клеток, которые также делятся неограниченно долго.
Однако при делении стволовых клеток одна из дочерних клеток может пойти по пути дифференцировки (то есть превратиться в обычную смертную клетку с определенными функциями: клетку легких, печени и т.п.), а другая — остаться "бессмертной". Поэтому стволовые клетки служат постоянным источником разнообразных клеток организма.
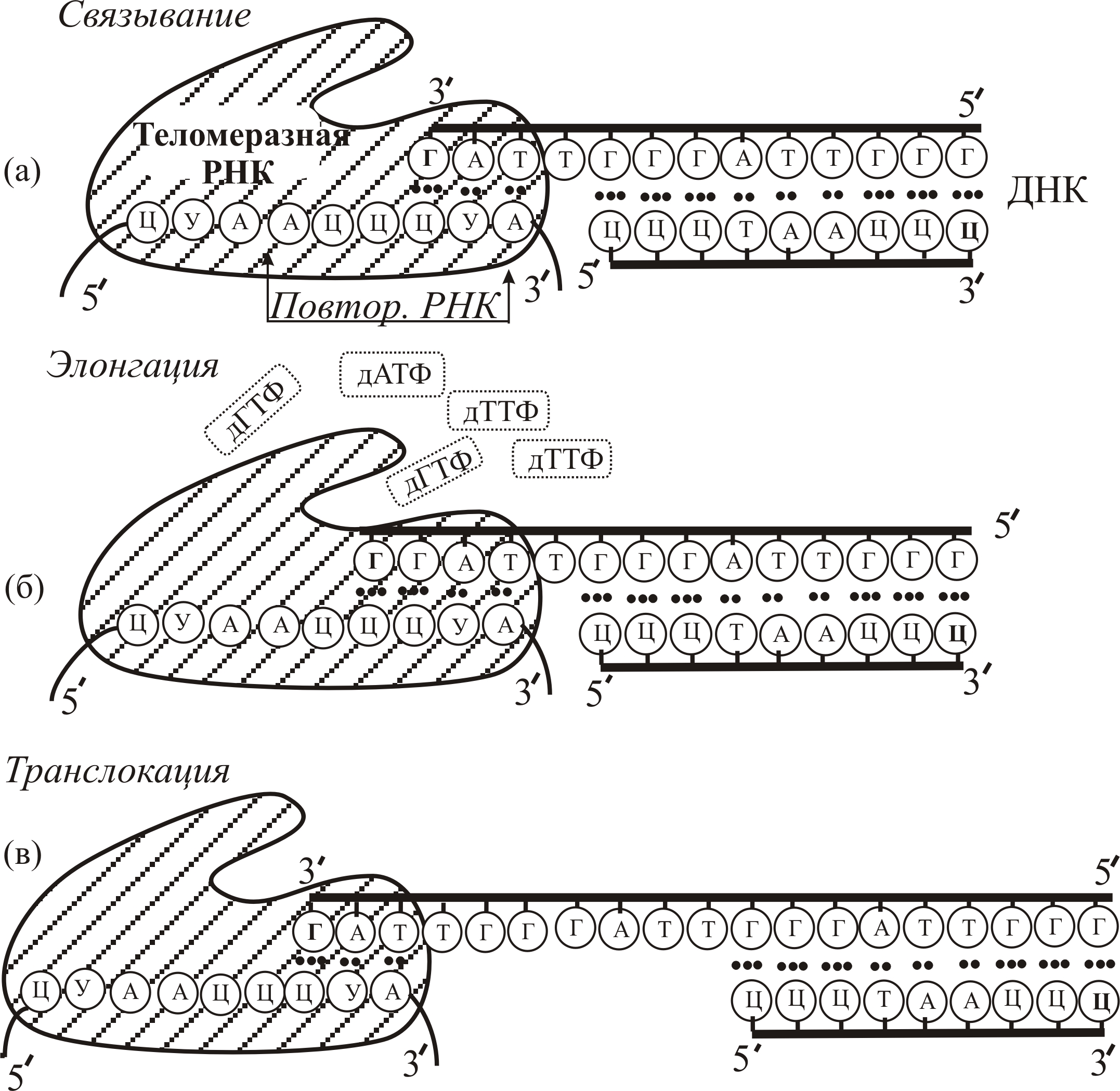
Рис. 9–28. Схема матричного синтеза одного теломерного ДНК-повтора с помощью теломеразной РНК: (а) – связывание теломеразы с 3´- концом хромосомной ДНК; (б) – элонгация, то есть присоединение субстратов-мономеров: дАТФ – дезоксиаденозин-5´-трифосфата, дГТФ – дезоксигуанозин-5´-трифосфата и дТТФ – дезокситимидин-5´-трифосфата; (в) – транслокация, то есть перемещение теломеразы для синтеза следующего повтора. Маленькие черные точки обозначают водородные (двойные или тройные) связи между соответствующими комплементарными основаниями соседних цепей