

Быстрые тепловые конформационные переходы
Для описания кинетики динамической подвижности белков можно использовать модифицированную модель диффузионного движения молекул в жидкости (см. ч.III, с. 215–217). Напомним, что диффузионное движение молекулы в жидкости складывается из ее колебательного (с частотой 0) движения в потенциальной яме, создаваемой молекулами, окружающими данную молекулу, и перескоков с характерной частотой п из одного равновесного положения в другое – образующуюся рядом флуктуационную полость.
Однако, в отличие от жидкостей диффузионное движение жестких фрагментов белков в вязкой среде имеет ряд принципиальных особенностей.
1. Атомы в каждом фрагменте ковалентно связаны и совершают высокочастотные колебания, соответствующие внутренним степеням свободы, с малыми амплитудами, как в обычных молекулах. Поскольку пространственная структура белка гетерогенна, то различные фрагменты обладают различными подвижностями, отличающимися характерными временами и амплитудами смещений. При переходе белковой глобулы из одного конформационного состояния в другое, фрагменты белка смещаются в определенных направлениях на расстояния порядка 0,1 нм. Смещения каждого фрагмента принято описывать конформационной координатой х. Так как предельная величина смещения ограничена, то фрагменты при конформационном переходе смещаются лишь в ограниченной области пространства.
Фрагменты в цепи белка связаны ковалентными связями и поэтому не могут перемещаться независимо. Смещение отдельного фрагмента в направлении х вызывает деформацию цепи и возникновение упругой силы Fупр(x), стремящейся вернуть фрагмент в первоначальное положение. Поскольку всякая сила связана с изменением потенциальной энергии системы, то удобно ввести конформационный потенциал
 ,
связанный с силой Fупр(x)
соотношением:
,
связанный с силой Fупр(x)
соотношением:
![]() . (3.15)
. (3.15)
Конформационный потенциал описывает форму потенциальной ямы для данного фрагмента, в которой он совершает ограниченное движение при переходе белка из одной конформации в другую. В общем виде зависимость имеет вид:
![]() . (3.16)
. (3.16)
Для квадратичной зависимости U(x)
коэффициент
![]() .
.
3. Если бы фрагмент мог свободно
перемещаться в пространстве, то переход
из состояния с конформационной координатой
(![]() )
в состояние с координатой (
)
в состояние с координатой (![]() )
совершался бы, при соответствующей
температуре, по горизонтальной прямой
(см. рис. 3–91). Но,
поскольку среднее расстояние между
фрагментами в белковой молекуле и
амплитуда колебаний атомов внутри
фрагментов существенно меньше
)
совершался бы, при соответствующей
температуре, по горизонтальной прямой
(см. рис. 3–91). Но,
поскольку среднее расстояние между
фрагментами в белковой молекуле и
амплитуда колебаний атомов внутри
фрагментов существенно меньше
![]() ,
то переход по прямой линии невозможен.
,
то переход по прямой линии невозможен.
Чтобы некоторый фрагмент белка мог
переместиться как целое на расстояние
![]() нм (превышающее амплитуду колебаний
составляющих его атомов (0,01÷0,1)Å), рядом
должна образоваться полость в результате
флуктуационных смещений молекулярных
групп, окружающих данный фрагмент, то
есть перемещение фрагмента имеет
диффузионный характер. Поэтому
реально переход фрагмента между
состояниями
и
может осуществляться путем серии
термически активированных перескоков
между локальными потенциальными ямами,
в которых фрагменты белка совершают
колебательное движение с собственными
частотами
нм (превышающее амплитуду колебаний
составляющих его атомов (0,01÷0,1)Å), рядом
должна образоваться полость в результате
флуктуационных смещений молекулярных
групп, окружающих данный фрагмент, то
есть перемещение фрагмента имеет
диффузионный характер. Поэтому
реально переход фрагмента между
состояниями
и
может осуществляться путем серии
термически активированных перескоков
между локальными потенциальными ямами,
в которых фрагменты белка совершают
колебательное движение с собственными
частотами
![]() в течение времени между двумя
последовательными перескоками. Прыжковая
диффузия по локальным конформационным
подсостояниям (обозначенными
светлыми точками) иллюстрируется
рис. 3–91.
в течение времени между двумя
последовательными перескоками. Прыжковая
диффузия по локальным конформационным
подсостояниям (обозначенными
светлыми точками) иллюстрируется
рис. 3–91.
Чтобы переместиться из одного конформационного подсостояния в другое, фрагмент должен израсходовать часть своей кинетической энергии для смещения соседних групп. Следовательно, для движения фрагмента требуется энергия активации Еа. Таким образом, движение фрагмента представляет сбой последовательное преодоление локальных потенциальных барьеров.
|
Рис. 3–91. Профиль потенциальной энергии U(x) (сплошная линия). Флуктуации энергии (пунктир) приводят к образованию локальных минимумов и барьеров, высота Еа и положение которых флуктуируют за счет взаимодействия с другими степенями свободы. ха – среднеквадратичная амплитуда ограниченной диффузии при температуре Т |
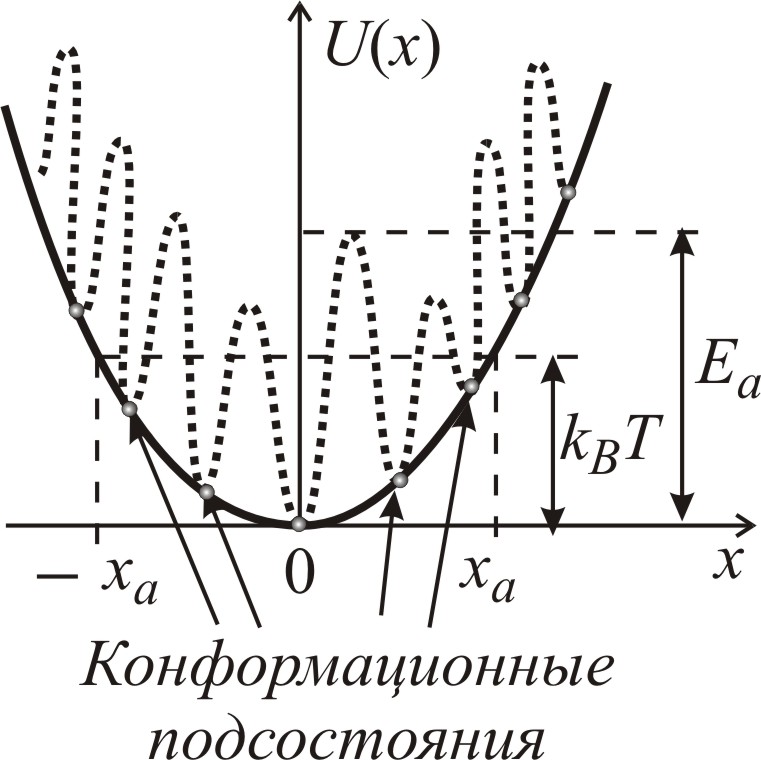
Отсюда следует, что движение фрагмента
характеризуется двумя пространственными
масштабами. Во-первых, – микросмещениями
между конформационными подсостояниями,
происходящими при благоприятном
перемещении соседних групп белка. Это
диффузионное движение характеризуется
коэффициентом конформационной диффузии
D(x),
зависящим от конфигурации окружающей
среды. Здесь х – конформационная
координата, указывающая положение
системы на пути конформационных
преобразований. Второй пространственный
масштаб (![]() )
характеризует существенно более
медленную перестройку конформации
белковой молекулы при переходе из одного
биологически активного состояния (
)
в другое (
).
Такой перестройке соответствует движение
всей системы вдоль конформационной
координаты – оси реакции – по
профилю плавно изменяющегося
конформационного потенциала U(x)
(рис. 3–91).
)
характеризует существенно более
медленную перестройку конформации
белковой молекулы при переходе из одного
биологически активного состояния (
)
в другое (
).
Такой перестройке соответствует движение
всей системы вдоль конформационной
координаты – оси реакции – по
профилю плавно изменяющегося
конформационного потенциала U(x)
(рис. 3–91).
4. Учтем теперь, что конформационные
перестройки белковых макромолекул
происходят в жидкой среде, которую можно
характеризовать микровязкостью
![]() .
При движении фрагментов в жидкой среде
на них действует сила вязкого трения
.
При движении фрагментов в жидкой среде
на них действует сила вязкого трения
![]() , (3.17)
, (3.17)
где
![]() – коэффициент силы вязкого трения,
– коэффициент силы вязкого трения,
![]() - скорость движения фрагментов.
Коэффициенты
и
связаны соотношением Стокса
- скорость движения фрагментов.
Коэффициенты
и
связаны соотношением Стокса
![]() , (3.18)
, (3.18)
где
![]() – эффективный линейный размер фрагмента
(1÷10)Å.
– эффективный линейный размер фрагмента
(1÷10)Å.
Кроме силы терния на фрагменты действует
сила
![]() ,
обусловленная столкновениями фрагментов
с молекулами жидкости, как при броуновском
движении:
,
обусловленная столкновениями фрагментов
с молекулами жидкости, как при броуновском
движении:
![]() , (3.19)
, (3.19)
где
![]() – случайные моменты времени. Амплитуда
силы
– случайные моменты времени. Амплитуда
силы
![]() также является случайной величиной.
также является случайной величиной.
С учетом всех, действующих на фрагмент сил, движение фрагмента описывается уравнением Ланжавена (M – масса фрагмента):
![]() .
(3.20)
.
(3.20)
Это стохастическое (вероятностное)
уравнение, требующее задания
функции распределения
.
Его решением также являются функции
распределения:
![]() и
и
![]() .
.
Если считать, что для перехода макромолекулы из одного конформационного состояния в другое отдельные фрагменты должны сместиться в среднем на расстояние х0, то зависимость от времени среднего квадратичного смещения фрагментов можно приближенно описать формулой
![]() . (3.21)
. (3.21)
Здесь
![]() – среднее квадратичное смещение,
необходимое для перестройки белка,
– среднее квадратичное смещение,
необходимое для перестройки белка,
![]() – время релаксации, определяющееся
величиной коэффициента трения или
микровязкостью среды:
– время релаксации, определяющееся
величиной коэффициента трения или
микровязкостью среды:
![]() ,
(3.22)
,
(3.22)
Из (2) следует, что при t = 0
смещение равно нулю
![]() ,
а при
,
а при
![]() смещение стремится к среднему значению
смещение стремится к среднему значению
![]() .
.
Поскольку микровязкость белка экспоненциально зависит от температуры
![]() , (3.23)
, (3.23)
где Еа – энергия активации вязкого движения), то время релаксации экспоненциально уменьшается при повышении температуры. Соответственно скорость диффузии при повышении температуры увеличивается, и, наоборот, при понижении температуры – замедляется.
Для белков при Т = 300 К
коэффициент вязкости составляет
![]() ,
что много больше вязкости воды (
,
что много больше вязкости воды (![]() ),
средней вязкости биологической мембраны
(0,1÷1)Па∙с и вязкости глицерина (
),
средней вязкости биологической мембраны
(0,1÷1)Па∙с и вязкости глицерина (![]() );
энергия активации
);
энергия активации
![]() ;
среднеквадратичная амплитуда
смещения
;
среднеквадратичная амплитуда
смещения
![]() .
.
Элементы вторичной структуры белков ( и ) совершают ограниченное диффузионное движение со средней амплитудой, зависящей от их массы и микровязкости среды. Амплитуда изгибных флуктуаций -спиралей достигает нескольких ангстрем.