

Этапы эмбрионального развития и дифференцировки.
Таблица № 2
Периоды собственно эмбрионального развития |
1 ———— Периоды эмбриональной дифференцировки |
I. Зигота - zygota до 30 часов |
I. Презумптивная - оотипическая Ovum - яйцеклетка, зигота |
II. Дробление - fissio До конца 7,5 суток |
II. Бластомерная - blastomerulis до конца 7,5 суток |
III. Гаструляция - gastrulatio от 7,5 суток до 21 суток |
III. Зачатковая - germinalia до 21 суток |
IV. Гистогенез - histogenesis Органогенез - organogenesis С 21 суток до рождения |
IV Тканевая, органная, организменная - до рождения гистогенез, органогенез, системогенез |
Выделяют два вида дифференцировки - эмбриональную и постэмбриональную, каждая из которых происходит поэтапно и соответствует определенным стадиям собственно эмбриогенеза.
Дифференцировка распространяется на все уровни организации материи: за счет цитологической дифференцировки обеспечивается разнообразие клеток; гистологической - образование тканей, а за счет половой дифференцировки - различие между женским и мужским организмом. Клеточная дифференцировка является одной из актуальных проблем биологии и медицины. Именно в ходе дифференцировки осуществляются сложные взаимосвязанные процессы, на основании которых происходят все изменения как в эмбриональном, так и в постэмбриональном онтогенезе.
Соответствие этапов собственно эмбрионального развития человека и этапов дифференцировки можно проследить в таблице № 2:
Прогенез.
Началу индивидуального развития нового организма предшествует возникновение и развитие в родительских организмах половых клеток - гамет, отличающихся друг от друга по строению и тем функциональным задачам, которые перед ними ставит природа.
Развитие мужских половых клеток сперматозоидов называется сперматогенезом и происходит в мужских половых железах - семенниках.
Развитие женских половых клеток называется овогенезом и происходит в женских половых железах - яичниках.
Одним из основных условий, необходимым для полового размножения, является сохранение из поколения в поколение постоянного числа хромосом. Это достигается путем уменьшения - редукции в половых клетках числа хромосом которое из диплоидного (46) становится гаплоидным (23). Затем при слиянии половых клеток во время оплодотворения диплоидный набор хромосом восстанавливается. Уменьшение хромосом в гаметах достигается мейозом, особым способом деления, характерным только для половых клеток. Мейоз состоит из двух, следующих друг за другом, митотических делений, причем перед вторым делением мейоза удвоения молекул ДНК не происходит.
Сущность мейоза состоит в том, чтобы каждая гамета получила гаплоидный набор хромосом.
В отличие от митоза, продолжительность которого обычно измеряется в минутах или часах, мейоз очень растянут во времени и занимает у мужчин до 72 дней, а у женщин нескольких дней до 40-50 лет.
Принципиально весь гаметогенез, как овогенез, так и сперматогенез, состоит из нескольких периодов.
Первые три периода аналогичны по характеру происходящих изменений для мужских и женских половых клеток. Это следующие периоды:
Период размножения.
Период роста.
Период созревания.
Для овогенеза развитие заканчивается периодом созревания и образованием овоцита второго порядка, который при оплодотворении даст зрелую яйцеклетку. А для сперматогенеза существует еще четвертый период - период формирования, во время которого происходят изменения в строении клетки, в результате чего и образуется узко специализированная, подвижная жгутиковая клетка - сперматозоид.
Выделяют следующие общие черты гаметогенеза, характерные для мужских и женских половых клеток.
1. Период размножения - заключается в том, что гаметы или стволовые половые клетки многократно делятся путем митоза и образуют необходимый резерв сперматогоний в мужском или овогоний в женском организме. Каждая из этих клеток
содержит диплоидный набор хромосом (46 хромосом): 44-аутосомы и 2-половые хромосомы: в мужском организме - XY, в женском - XX.
Период роста. В него постепенно вступают отдельные сперматогонии и овогонии, в которых происходит первоначальная дифференцировка. Увеличиваются размеры клеток, и они начинают готовиться к делению. На этой стадии клетки и получают название сперматоциты 1 порядка и овоциты 1 порядка. Профаза 1 деления - длительная, сложная стадия, в которой обычно выделяют еще 5 подстадий: лептотенную, зиготенную, пахитенную, диплотенную и диакинез.
Лептотенная стадия. Хромосомы имеют форму тонких (leptos - тонкий), длинных нитей и состоят из двух хроматид. Но уже с этой стадии начинается процесс спирализации хромосом.
Зиготенная стадия. Гомологичные хромосомы сближаются и располагаются одна подле другой, образуя 23 пары хромосом: 22 пары аутосом и 1 пара половых хромосом. Такое близкое расположение хромосом в паре называется коньюгацией. Продолжается спирализация и укорочение хромосом.
В начале пахитенной стадии завершается конъюгация хромосом и две рядом лежащие хромосомы образуют бивалент ши диаду. Для этой стадии за счет спирализации характерно еще большее утолщение и укорочение хромосом. В конце пахитенной и начале диплотенной стадии происходит кроссинговер, когда гомологичные хромосомы обмениваются генетическим материалом. Кроссинговер является одним из основных механизмов возникновения генетических различий между потомками.
После завершения кроссинговера начинается диплотенная стадия, особенностью которой является процесс расщепления хромосом на составляющие их хроматиды, что приводит к удвоению хромосом. Отделение хроматид происходит практически по всей длине хромосомы, связанными они остаются лишь в одной точке. Таким образом, каждый бивалент состоит из 4-хроматид и теперь его называют тетрадой.
На стадии диакинеза хромосомы еще больше укорачиваются, поэтому четче обозначается каждая хроматида в 23 тетрадах. К этому времени ядрышко в ядре сперматоцита первого порядка и овоцита первого порядка исчезает, ядерная мембрана фрагментируется, в это же время и заканчивается формирование веретена деления.
В период созревания происходят метафаза, анафаза и телофаза первого деления мейоза.
В метафазу первого деления, когда тетрады хромосом выстраиваются по экватору клетки и связываются с нитями веретена. Каждая пара хромосом укладывается по экватору клетки так, что одна хромосома лежит по одну сторону экватора, а другая - по другую, т.е. хромосомы материнского и отцовского происхождения располагаются по обе стороны экватора случайным образом. Поэтому при последующих стадиях деления в одну клетку может попасть больше материнских, а в другую - отцовских хромосом. Такое случайное распределение хромосом - способствует созданию генетических различий у потомков.
Анафаза первого деления. На этой стадии гомологичные хромосомы в каждой паре отходят друг от друга к противоположным полюсам клетки, а так как в ядре содержится 23 пары хромосом, то к каждому полюсу отойдет по 23 хромосомы, каждая из которых состоит из 2 хроматид. Таким образом, особенностью первого деления мейоза является то, что к полюсам клетки уходят целые хромосомы из двух хроматид отцовского или материнского набора. Это приводит к образованию генетически разнородных клеток.
Телофаза первого деления. На полюсах клетки идет образование дочерних ядер, каждое из которых содержит диплоидное число хромосом.
Следовательно, при 1-м делении мейоза из одного сперматоцита 1 порядка образуются 2 идентичные клетки - сперматоциты 2 порядка. Особенностью является то, что из каждого овоцита 1 порядка формируется 2 неравнозначные клетки: одна крупная это овоцит 2 порядка и вторая, мелкая это 1-ое направительное тельце (редукционное).
![]()
Первое деление мейоза заканчивается цитотомией и образованием 2 новых клеток, но хромосомы в них остаются удвоенными, поэтому при подготовке ко второму делению мейоза нового удвоения ДНК не происходит, и после короткой интерфазы клетки вступают во второе деление мейоза
Второе деление мейоза протекает как митоз, с той лишь разницей, что в интерфазу не происходит удвоения ДНК, так как исходные клетки уже имеют диплоидное число хромосом, состоящих из 2 хроматид. В метафазу хроматиды располагаются по экватору, поэтому в анафазе к полюсам клетки расходятся по 23 хроматиды, или гаплоидный набор. В телофазу второе деление мейоза завершается.
Таким образом в результате мейоза каждая исходная половая клетка с диплоидным набором хромосом (сперматоцит 1 порядка и овоцит 1 порядка), вступив в мейоз и пройдя два деления, даст 4 клетки с гаплоидным набором хромосом. В мужском организме все 4 гаплоидных клетки равноценны и называются сперматидами. В женском организме, вследствие ассиметрии мейоза, образуется только одна половая клетка на стадии овоцита второго порядка, а три другие дочерние клетки дадут направительные (редукционные) тельца с гаплоидными ядрами. Овоцит второго порядка находится в просвете маточной трубы на стадии метафазы второго деления созревания. Для завершения овогенеза и образования яйцеклетки необходимо проникновение в овоцит II порядка сперматозоида.
Таким образом, несмотря на сходство основных процессов сперматогенеза и овогенеза, между ними существуют значительные различия, которые хорошо прослеживаются в таблице № 3.
Прогенез. Общие и отличительные черты гаметогенеза женской и мужской половых
клеток
Таблица№ 3
|
Овогенез |
Сперматогенез |
|
Период размножения |
Митоз |
Овогонии в яичнике - со 2-го по 7-й мес. эмбриогенеза |
Сперматогонии - в семеннике, со 2-го по 7-й месяц эмбриогенеза, всю жизнь |
Период роста |
Профаза первого мейотического деления |
Овоциты I порядка в яичнике - с 7 мес. эмбриогенеза до овуляции в течение жизни |
Сперматоциты I порядка в семеннике, с рождения, всю жизнь. |
Период созревания |
Мейоз 1 деление |
Овоциты II порядка в воронке яйцевода, в течение 36 час. После овуляции |
Сперматоциты II пор. - в семеннике, с 6 лет всю жизнь. |
Мейоз 2 деление |
Яйцеклетка в воронке яйцевода, после оплодотворения (дифференцировка завершена). |
Сперматиды - в семеннике, с полового созревания всю жизнь. |
|
Период формирования |
Дифференцировка клеток |
Отсутствует. |
Сперматозоиды - в семеннике, с полового созревания всю жизнь. |
Особенности строения яйцеклетки.
В конце периода созревания в газетах совершается один из наиболее важных процессов дифференцировки - Презумптивная (предположительная) дифференцировка. Она приводит к образованию радикально отличающихся типов клеток - яйцеклетки и сперматозоида.
Яйцеклетка является самой крупной клеткой в организме, ее диаметр колеблется от 120 - до 200 мкм, а объем тысячекратно больше объема сперматозоидов. С поверхности она окружена 2-3 тысячами фолликулярных клеток, которые формируют лучистый венец и зернистую оболочку. Регулирующую роль в период презумптивной оотипической дифференцировки играет ядро, изменения которого способствую!' реализации программы сегрегации цитоплазмы. Сегрегацией называется жесткое закономерное перемещение и расположение органоидов и химических веществ по определенным участкам тела клетки Основным процессом, происходящим в ядре, является амплификация на молекуле ДИК генов. Термин «амплификация» обозначает копирование или увеличение количества. Поэтому, в результате амплификации происходит резкое увеличение молекул ДНК и РНК, которые будут участвовать в синтезе белков в клетках эмбриона, а не самой яйцеклетки. Яйцеклетка запасает такое количество РНК всех типов (рРНК, иРНК, тРНК), которого хватает для осуществления синтеза белка во всех клетках на первых трех этапах эмбрионального развития (зигота, дробление и гаструляция). Без амплификации синтез РНК в новых клетках эмбриона растянулся бы на длительный срок, например у шпорцевой лягушки - на 500 лет, а у человека - на десятилетия.
 Участки
ДНК с амплифицированными кодонами
замыкаются в кольца, перемещаются к
внутреннему листку ядерной оболочки и
вступают с ним в контакт. В дальнейшем,
этот участок оболочки начинает
выпячиваться в цитоплазму, отпочковывается
и становится частью цитоплазмы. Так
создается новый, характерный только
для овоцита, органоид - пористая мембрана
(см. рис.№3). Пористая мембрана представляет
собой регуляторный комплекс, предназначенный
для синтеза белка в
новых клетках эмбриона.
Участки
ДНК с амплифицированными кодонами
замыкаются в кольца, перемещаются к
внутреннему листку ядерной оболочки и
вступают с ним в контакт. В дальнейшем,
этот участок оболочки начинает
выпячиваться в цитоплазму, отпочковывается
и становится частью цитоплазмы. Так
создается новый, характерный только
для овоцита, органоид - пористая мембрана
(см. рис.№3). Пористая мембрана представляет
собой регуляторный комплекс, предназначенный
для синтеза белка в
новых клетках эмбриона.
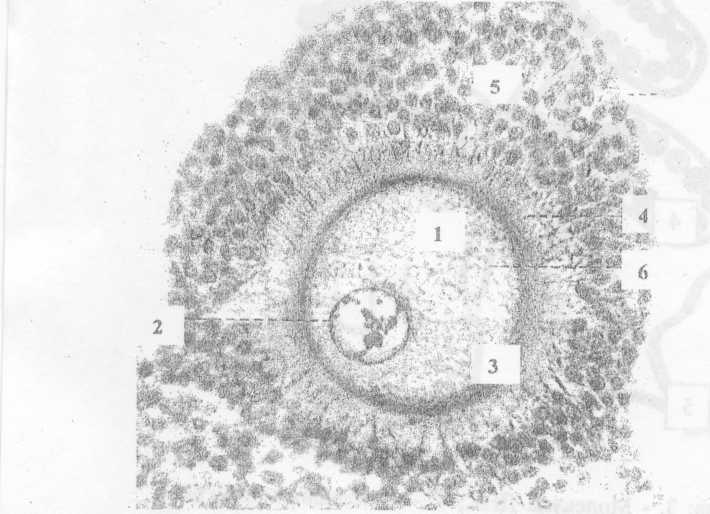
Яичник.
Окраска гематоксилин-эозином.
Рис.№1
3 - ядро оопита
- ядрышки.
- блестящая.
-лучистый венец и 5 - фолликулярные клетки.
- желточные гранулы - слоистые пластинки.
![]()
A i
ЭлектромограМма - яйцеклетка и фолликулярные клетки
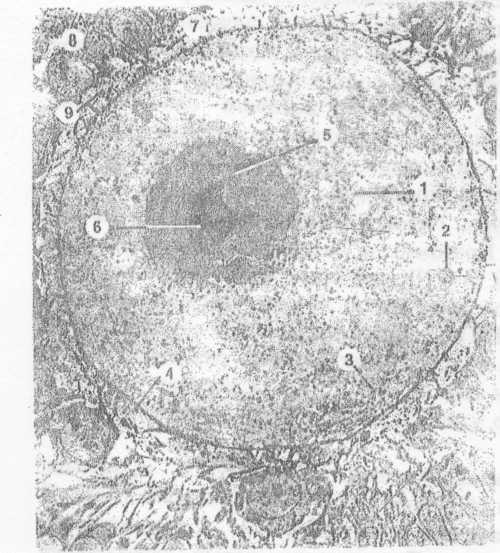 Рис.
№ 2
Рис.
№ 2
- желточные гранулы содержат фосфовитии и липовителлин;
- кортикальные гранулы; содержат
гидролитические ферменты,
участвующие в кортикальной
реакции;
- плазмолемма может образовывать микроворсинки;
- ядро ооцита;
- ядрышко;
- блестящая оболочка;
- фолликулярные клетки и их
отростки - 9 проникающие в
блестящую оболочку.
Процесс формирования органелл «мористых мембран»
РисЗ.
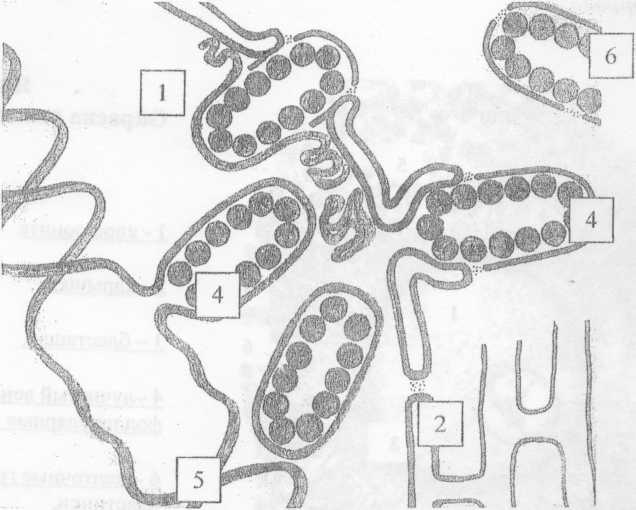
-Ядро; 2. - Ядерная мембрана; 3. - Молекула ДНК; 4. - Амплификация РНК; 5. - Выход в цитоплазму; 6. - Органоид пористая мембрана.
В ходе оотииической дифференцировки в результате сегрегации происходит функциональное разъединение цитоплазмы овоцита на 2 зоны:
© Центральную, лежащую вокруг ядра.
© Периферическую - кортикальную.
В центральной зоне расположены органоиды двух функциональных аппаратов клетки: синтетического и энергетического. Синтетический аппарат включает
Яйцеклетка является самой крупной клеткой в организме, ее диаметр колеблется от 120 - до 200 мкм, а объем тысячекратно больше объема сперматозоидов. С поверхности она окружена 2-3 тысячами фолликулярных клеток, которые формируют лучистый венец и зернистую оболочку. Регулирующую роль в период презумптивной оотипической дифференцировки играет ядро, изменения которого способствуют реализации программы сегрегации цитоплазмы. Сегрегацией называется жесткое закономерное перемещение и расположение органоидов и химических веществ по определенным участкам тела клетки Основным процессом, происходящим в ядре, является амплификация на молекуле ДНК генов. Термин «амплификация» обозначает копирование или увеличение количества. Поэтому, в результате амплификации происходит резкое увеличение молекул ДНК и РНК, которые будут участвовать в синтезе белков в клетках эмбриона, а не самой яйцеклетки. Яйцеклетка запасает такое количество РНК всех типов (рРНК, иРНК, тРНК), которого хватает для осуществления синтеза белка во всех клетках на первых трех этапах эмбрионального развития (зигота, дробление и гаструляция). Без амплификации синтез РНК в новых клетках эмбриона растянулся бы на длительный срок, например у шпорцевой лягушки - на 500 лет, а у человека - на десятилетия.
 Участки
ДНК с амшшфицированными кодонами
замыкаются в кольца, перемещаются к
внутреннему листку ядерной оболочки и
вступают с ним в контакт. В дальнейшем,
этот участок оболочки начинает
выпячиваться в цитоплазму, отпочковывается
и становится частью цитоплазмы. Так
создается новый, характерный только
для овоцига, органоид - пористая мембрана
(см. рис.№3). Пористая мембрана представляет
собой регуляторный комплекс, предназначенный
для синтеза белка в
новых клетках эмбриона.
Участки
ДНК с амшшфицированными кодонами
замыкаются в кольца, перемещаются к
внутреннему листку ядерной оболочки и
вступают с ним в контакт. В дальнейшем,
этот участок оболочки начинает
выпячиваться в цитоплазму, отпочковывается
и становится частью цитоплазмы. Так
создается новый, характерный только
для овоцига, органоид - пористая мембрана
(см. рис.№3). Пористая мембрана представляет
собой регуляторный комплекс, предназначенный
для синтеза белка в
новых клетках эмбриона.
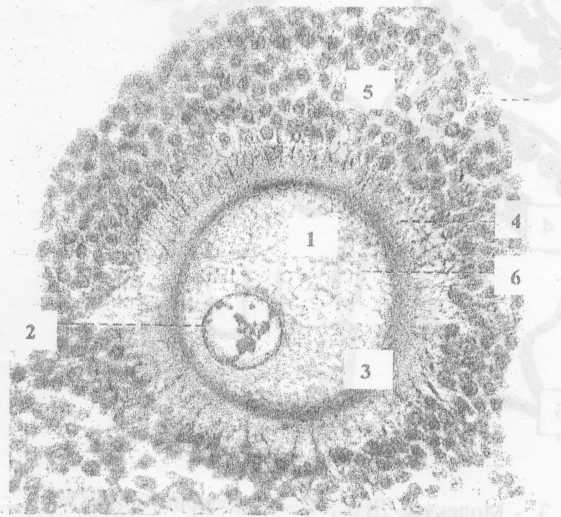
Яичник,
Окраска гематоксилин-эозином.
Рис.№1
- ядро ооцита
•• ядрышки.
- блестящая.
-лучистый венец и 5 - фолликулярные клетки,
- желточные гранулы - слоистые пластинки.
гранулярную ЭПС, комплекс Гольджи и пористые мембраны. Пористых мембран тысячи, они лежат между ЭПС и КГ.
Энергетический аппарат представлен огромным количеством крупных митохондрий, их в 12-17 раз больше, чем в соматических клетках. Митохондрии яйцеклетки имеют следующие особенности: 1) делятся почкованием и укладываются вокруг ядра в радиальном направлении; 2) 90% митохондрий в яйцеклетке неактивны, так как они создаются для новых клеток эмбриона, где и будут работать, 3) все митохондрии наследуются только по материнской линии.
Одним из факторов, определяющим диффереицировку эмбриона, является уникальный белковый состав цитоплазмы яйцеклетки и депонирование в ней специфических структур - слоистых пластинок. Установлено, что все белки яйцеклетки можно разделить на три группы:
Организация цитоплазмы клетки настолько сложна и уникальна, что созданы специальные карты, аналогичные географическим, отражающие закономерность расположения органоидов и включений в овоците (Де Робертис, 1964).
Схема структурной организации яйцеклетки
Рис 4.
![]()
А
r-
AS'
I Of
Ъ
0г
н
7
А'
-4-Т И
/
/V
к!
£ Г
ЗС
О
23
.
■ Л
ОБРАЗУЕМЫЕ
(“Б£Л
КИ ОБРАЗУЕМЫЕ
.
. ЕМЫЕ яйцеклеткой
в
ПЕЧЕНИ чо".
(ВИТЕ
Л АО - ХГ/
ГЕНИН)
ОБРАЗУЕМЫЕ
ФОЛЛИКУЛЯРНЫМИ
КЛЕТКАМИ ^
Х
V щщ
|| ВЕГЕТАТИВНЫЙ
полное
Л?
ЗЛ
Им
jw*
I
Лг, V =§>0










® Белки, синтезируемые самой яйцеклеткой.
» Белки, синтезируемые вспомогательными фолликулярными клетками половых желез.
« Белки, синтезируемые в печени матери.
Белки 1-ой группы являются регуляторами деления клеток и играют очень важную роль на втором этапе эмбриогенеза - дроблении. В цитоплазме яйцеклетки эти белки заблокированы и активируются ферментами сперматозоида только после оплодотворения. Белки 2-ой группы используются самой яйцеклеткой для пластических целей.
Белок 3-ей группы - вителлогенин, представляет собой сложный комплекс, который состоит из двух частей: липовителлина и фосфовитина. Оба белка формируют сложные структуры липопротеинового комплекса в виде слоистых пластинок. Каждая пластинка отграничена биомембраной и имеет кристаллическое строение центральной зоны. Этот комплекс упакованных биомембран будет использоваться новыми клетками па протяжении первых трех периодов эмбрионального развития. Если микрохирургическим путем удалить какую - либо часть яйцеклетки, то не разовьется определенная ткань или даже орган, гак как все части цитоплазмы строго распределены в результате презумптивной дифференцировки.
Периферическую часть цитоплазмы составляет узкая кортикальная зона шириной 2-3 мкм. Здесь накапливаются следующие структуры и вещества: неактивные лизосомы, регуляторы оплодотворения - гамоны, сложные углеводы, сократительные белки и гликопротеиды, которые лежат в примембранном слое. Количество лизосом достигает 5000 штук, они имеют крупные размеры, лежат вплотную к цитолемме и обуславливают неровность ее поверхности. Неактивные лизосомы называются кортикальными гранулами, которые в момент оплодотворения активируются и выбрасываются из яйцеклетки. Это так называемая кортикальная реакция, в результате которой ферменты лизосом способствуют инактивации сперматозоидов, окружающих уже оплодотворенную яйцеклетку.
Функции кортикальной зоны:
® Способствует оплодотворению и моногамии или проникновению одного спермия.
© Обеспечивает движение клеточной оболочки в период деления за счет сократительных белков.
Также во время кортикальной реакции происходит выделение гликопротеидов и гамонов за пределы клетки. Гликопротеиды участвуют в образовании гидратированной оболочки, которая превращается в прочную оболочку оплодотворения.
Особо сложную дифференцировку претерпевае т надмембранный слой цитолеммы яйцеклетки. Этот слой представляет собой гликокаликс толщина которого достигает 1 -
мкм, что в сотни раз больше, чем у соматических клеток. Поэтому этот слой хорошо различим в световом микроскопе и при окраске определяется как блестящая оболочка - Zona pellucida - ZP. В составе ZP гликопротеины, имеющие тонкофибриллярную структуру. Особое значение имеют две группы гликопротеинов, которые обозначаются как ZP - 3 и ZP - 2. Гликопротеид ZP - 3 является рецептором для сперматозоидов, в блестящей оболочке их насчитывается до 1012. Гликопротеид ZP-2, наоборот, препятствует проникновению сперматозоидов в яйцеклетку после оплодотворения, способствуя моноспермии.
Итак, зрелая яйцеклетка имеет жесткую дифференцировку цитоплазмы и специализированную цитолемму, содержащую рецепторы к сперматозоидам. Вокруг закончившей дифференцировку яйцеклетки располагаются 2 - 3 тыс. фолликулярных клеток, осуществляющих для нее трофическую функцию.
Структура сперматозоидов.
Сперматозоид относится к жгутиковым клеткам, в мужском организме он неподвижен и начинает самостоятельно двигаться только в женских половых путях. Спермий, как и яйцеклетка, имеет гаплоидный набор хромосом, что позволяет при слиянии с яйцеклеткой восстановить диплоидный генотип клеток.
Рис. 5.
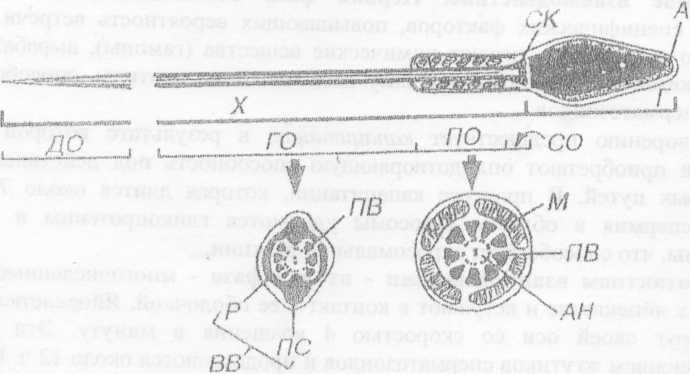
Строение
сперматозоида.
Половые хромосомы в клетках человека обозначают X и У. Для женского организма характерен набор XX, поэтому яйцеклетки в любом случае получают только половую хромосому X. В мужском организме набор ХУ, поэтому по половой хромосоме мужские половые клетки могут быть двух видов: одни - с X, другие - с У. Если яйцеклетка будет оплодотворена сперматозоидом, содержащим Х-хромосому, то будет развиваться девочка, а если сперматозоидом, содержащим Y-хромосому - мальчик. Следовательно, пол будущего ребенка определяется в момент оплодотворения спрематозоидом.
ДНК в сперматозоиде находится в сверхепирализованном кристаллическом состоянии. В клетке совершенно отсутствует РНК, поэтому никакие синтетические процессы не происходят. Из общих органоидов имеются 20 митохондрий, которые функционируют только в женских половых путях, видоизмененный клеточный центр, формирующий жгутик, видоизменений комплекс Гольджи - акросома, несущий лизирующие ферменты (см. рис.№ 5). Особое значение имеет фермент, регулирующий синтез ДНК. В спермии он накапливается в неактивном состоянии и активируется только после оплодотворения. Кроме того, в цитолемме спермия имеются 2 типа гормонов оплодотворения андрогамоны.
1.3. Оплодотворение и образование зиготы
Оплодотворением (fertilise! Lio) называется слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, и возникает качественно новая клетка - зигота (оплодотворенная яйцеклетка, или одноклеточный зародыш).
Для обеспечения оплодотворения общее количество сперматозоидов в сперме должно быть не менее 150 млн., а концентрация их в 1 мл -- не менее 60 млн. Хотя в яйцеклетку проникает только один из них, остальные спермии создают условия для оплодотворения. В половых путях женщины их число значительно уменьшается по направлению от влагалища к дистальному концу маточной трубы.
В процессе оплодотворения различают три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) вхождение
сперматозоида в яйцеклетку и последующее слияние ядер - сиигамия. Основные процессы, происходящие при оплодотворении, представлены на рис № б
Дистантное взаимодействие. Первая фаза обеспечивается хемотаксисом, совокупностью специфических факторов, повышающих вероятность встречи половых клеток. Важную роль в этом играют химические вещества (гамоны), вырабатываемые половыми клетками. Яйцеклетки в эту фазу выделяют еще пептиды, способствующие привлечению сперматозоидов.
Оплодотворению предшествует капацитация, в результате которой спермии активируются и приобретают ошюдотворя ющую способность под действием секрета женских половх>1х путей. В процессе капацитации, которая длится около 7 часов, с плазмолеммы спермия в области акросомы удаляются гликопротеины и протеины семенной плазмы, что способствует акросомалыюй реакции.
При контактном взаимодействии - вторая фаза ~ многочисленные спермии приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает вращаться вокруг своей оси со скоростью 4 вращения в минуту. Эти движения обусловлены биением жгутиков сперматозоидов и продолжаются около 12 ч. В эту фазу спермиях происходит акросомальная реакция, в результате которой наружная мембрана акросомы сливается с плазмолеммой спермия. Затем в области слияния мембраны разрываются, ферменты акросомы выходят в окружающую среду, и блестящая зона оказывается в непосредственном контакте с акросомальными ферментами. Ферменты разрушают блестящую зону, поэтому спермий проходит через нее и входит в пери вителлин овое пространство, расположенное между блестящей зоной и плазмолеммой яйцеклетки.
Схема процесса оплодотворения (но Вассерману с изменениями).
Рис № 6.
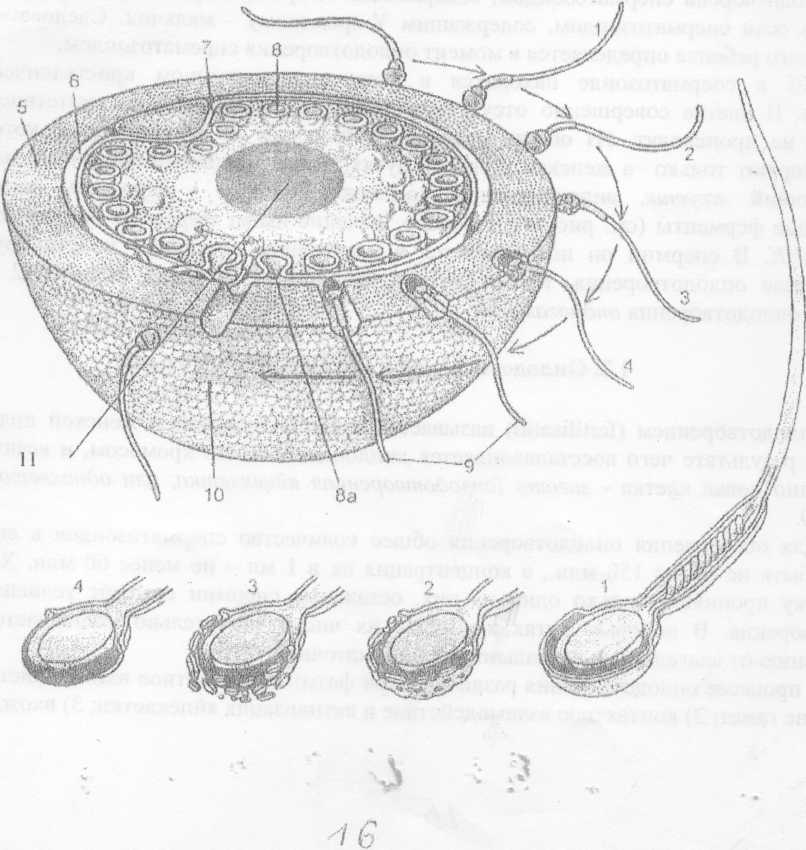
2, 3, 4 - стадии акросомной реакции; 5 — zona pellucicla (блестящая зона); 6 - перивителлиновое пространство; 1 - направительное тельце, 8 - кортикальная гранула; 8а
кортикальная реакция; 9 - вхождение спермия в яйцеклетку; 10 - зонная реакция, 11. - ядро яйцеклетки.
Только этот сперматозоид связывается с рецептором в плазмолеммс яйцеклетки - ZP-3R и может проникнуть в нее для оплодотворения. Через несколько секунд свойства плазмолеммы яйцеклетки изменяются, что запускает кортикальную реакцию. Во время кортикальной реакции плазмолеммы яйцеклетки сливается с мембранами кортикальных гранул, в результате чего содержимое гранул выбрасывается, и ферменты этих гранул воздействуют на гликопротеиды блестящей зоны, превращая ее в оболочку оплодотворения.
Таким образом, у млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Сотни тысяч других мужских половых клеток способствуют оплодотворению, так как выделяют ферменты (трипсин, гиалуронидаза), которые разрушают фолликулярные клетки лучистого венца и расщепляют глшсозаминогликаны прозрачной зоны яйцеклетки. Отделяющиеся фолликулярные клетки склеиваются в конгломерат, который вслед за яйцеклеткой перемещается в матку благодаря мерцанию ресничек эпителиальных клеток слизистой оболочки и затем в ней фагоцитируется.
Третья фаза. В яйцеклетку проникают головка и промежуточная часть хвостового отдела. После вхождения сперматозоида в яйцеклетку развивается зонная реакция. Вследствие зонной реакции молекулы - ZP3 модифицируются и утрачивают способность быть рецепторами спермиев. Параллельно происходит превращение ядер гамет в пронуклеусы, которые начинают сближаться.
Благодаря механизмам предотвращения полиспермии только одно гаплоидное ядро сперматозоида получает возможность слиться с гаплоидным ядром яйцеклетки, что приводит к восстановлению характерного для всех клеток диплоидного набора.
Взаимодействие сперматозоидов с яйцеклеткой может быть заблокировано при помощи антител против веществ, входящих в прозрачную зону. На этом основании изыскиваются способы иммунологической контрацепции.