6.3.2.2. Практическое применение теории
Предпринималось много попыток оценить реальный генетический груз в популяциях человека. Ранние работы представляли собой практическое приложение теории генетического груза. Однако более поздние исследования основаны на непосредственном использовании медицинрких данных.
354 6. Популяционная генетика
Таблица 6.20. Результаты нескольких исследований влияния инбридинга на смертность в младенчестве, детстве и юношеском возрасте в населении Японии |
|||||
Автор, регион |
А |
В |
В/А |
Размер инбредной выборки |
Метод определения |
|
|
|
|
|
|
Ватанабе, префектура Фукусима |
0,0881 |
0,5157 |
5,8 |
4 594 |
Учитывается смертность среди детей школьного возраста |
Танака, Кисимото, Сидзуока |
0,1253 |
0,7191 |
5,7 |
2205 |
Учитывается смертность среди детей школьного возраста |
Шелл и др., префектура Нагасаки (Куросима) |
0,0927 |
1,4074 |
15,2 |
223 |
Данные приблизительно за 15 лет, учитывается смертность до 20летнего возраста |
Шелл и Нилл, префектура Хиросима, префектура |
0,0875 |
0,5317 |
6,1 |
1697 |
Регистрация беременности на 5-м месяце; учитывается смертность |
Нагасаки |
0,0986 |
0,1060 |
1,1 |
2608 |
до 8-летнего возраста |
Шелл и Нил, Куре |
0,0929 |
0,0405 |
0,4 |
564 |
Регистрация беременности на 5-м месяце; учитывается смертность до 15-летнего возраста |
Янасе, префектура Фукуока (3 локальности) |
0,0962 |
1,2535 |
13,0 |
277 |
Посемейное обследование, учитывается смертность до 6 лет |
|
0,1292 |
0,3308 |
2,6 |
304 |
|
|
0,0916 |
0,9884 |
10,8 |
301 |
|
Фудзики и др., префектура Ямагути (3 локаль- |
|
|
|
|
Регистрационные книги и посемейный опрос; смертность до средне- |
ности) |
0,1222 |
0,3287 |
2,7 |
497 |
го детского возраста |
|
0,1985 |
-0,8107 |
-4,1 |
234 |
|
|
0,1936 |
-0,9608 |
-5,0 |
79 |
|
Нагано, префектура |
|
|
|
|
Учитывается смертность до 12-лет- |
Фукуока (гор. Фукуока) |
0,0873 |
0,6765 |
7,8 |
5 953 |
него возраста |
Шелл и др., префектура |
|
|
|
|
Посемейное обследование; смерт- |
Нагасаки (Хирадо) |
0,1157 |
0,7703 |
6,7 |
6626 |
ность от неслучайных причин в основном до 20-летнего возраста |
Фрейре-Майо и др., |
|
|
|
|
Посемейное обследование, смерт- |
японцы, иммигрировавшие в Бауру, шт. Сан-Паулу, Бразилия |
0,1378 |
0,6995 |
5,1 |
105 |
ность |
Среднее |
0,1036 |
0,6700 |
6,7 |
— |
— |
Эти работы будут рассмотрены в разд. 6.3.2.4.
Попытки оценить генетический груз путем анализа кровнородственных браков. Влияние кровного родства родителей на частоту мертворождений и детскую смертность исследовалось во многих работах. Наиболее полные и наиболее надежные во многих отношениях данные относятся к населению Японии [198; 1839; 1840; 1876].
В таблице 6.20 обобщаются результаты этих работ до 1972 г. Размеры контрольных инбредных выборок не приводятся; как правило, они превышают размеры инбредных выборок. Отношение В/А изменяется от +15,2 до — 5. Отрицательное значение В/А означает, что детская смертность в кровнородственных браках была ниже, чем в некровнородственных. Формально это означает наличие «отрицательного генетического груза», что с биологической точки зрения бессмысленно.
В большинстве работ, исследующих влияние кровного родства на генетический груз, получены оценки В/А, варьирующие между 5,7 и 7,8; простое усреднение дает отношение В/А, равное 6,7. Этот результат, вероятно, завышен благодаря более низкому социально-экономическому статусу
6. Популяционная генетика 355
Таблица 6.21. Влияние инбридинга и смертность [1763] |
|||
Литературный источник |
Исследованная популяция |
В/А (сравнение браков двоюродных сибсов с неродственными браками) |
|
|
Раса |
Регион |
|
Нил (1963)1' |
Негры |
Бразилия |
6,9 |
Нил (1963)" |
« |
« |
7,6 |
Фрейре-Майо (1963) |
« |
« |
9 |
Фрейре-Майо и Азеведо (1971) |
« |
« |
3 |
Нил (1963) |
« |
Танганьика |
-1,0 |
Нил (1963) |
Белые |
Бразилия |
-0,0 |
Нил (1963) |
« |
« |
-0,6 |
Фрейре-Майо (1963) |
« |
« |
1,0 |
Фрейре-Майо и Азеведо (1971) |
« |
« |
3 |
Фрейре-Майо и др. (1963) |
Смешанная |
« |
16,5 |
Нил (1963) |
Белые |
США |
7,2 |
Нил (1963) |
« |
Чикаго, Франция |
6,6 |
Нил (1963) |
« |
Морбиан, Франция |
20,2 |
Нил (1963) |
« |
Луар и Шер, Франция |
13,1 |
Нил (1963) |
« |
Сев. Швеция |
-3,0 |
Кумар и др. (1967) |
|
Керала |
20 |
Роберте (1969) |
Индийцы |
« |
14,8 |
1) Работа Нила суммирует данные многих авторов. Робертс модифицировал результаты Кумара для учета других степеней кровного родства, кроме двоюродных сибсов. |
|||
пар, вступивших в кровнородственный брак. Несмотря на то что в Японии кровнородственные браки социально намного больше распространены, чем в других странах, особенно с христианской религией, социально-экономические отклонения наблюдаются и там, причем эти отклонения имеют различные направления.
В исследованиях, проведенных в Южной Америке, Франции, США, Индии и Африке, этот эффект проявляется еще ярче (табл. 6.21). Для удобства сравнения оценка отношения В/А здесь ограничена браками между двоюродными братьями и сестрами, поэтому эти данные нельзя непосредственно сравнивать с данными табл. 6.20, составленной для населения Японии. Однако изменчивость здесь, вероятно, еще выше, чем между данными, полученными в работах японских авторов. Это неудивительно, поскольку в Европе и в США кровнородственные браки очень редки (табл. 6.18). В странах с сильными христианскими традициями общественное мнение выступает против таких браков и, как было показано выше, пары, вступившие в кровнородственный брак, отличаются от обычного населения в социальном и даже в психологическом отношении.
Один из способов устранения, по крайней мере частично, этих отклонений - использование в качестве контроля детей братьев и сестер лиц, вступивших в кровнородственный брак. В работе, в которой таким образом изучалось население Вогезских гор (Франция) [1768], было проведено сравнение 189 кровнородственных и 646 «контрольных» браков. Разница в уровне перинатальной смертности оказалась невелика, статистически недостоверна для браков двоюродных сибсов и пренебрежимо мала для браков, где между супругами была более отдаленная степень родства (табл. 6.22). Разница в числе стерильных браков между кровнородственными и некровнородственными браками была статистически значима. Этот результат, возможно (но не обязательно), указы-
356 6. Популяционная генетика
Таблица 6.22. Эффект инбридинга [103] |
||||||
|
Браки двоюродных сибсов |
Браки троюродных сибсов |
Все браки |
|||
|
Кровнородственные браки |
Контроль |
Кровнородственные браки |
Контроль |
Кровнородственные браки |
Контроль |
Среднее число детей на семью |
4,2 |
5,1 |
4,8 |
4,8 |
4,5 |
4,8 |
Процент стерильных браков |
— |
– |
– |
– |
6,9 |
4,6 |
Перинатальная смертность (%) |
11,1 |
9,0 |
8,0 |
7,9 |
8,9 |
8,5 |
вает на более высокую внутриутробную смертность детей от кровнородственных браков. Кроме того, необходимо учитывать, что в отношении В/А величина А отражает не только генетическую компоненту смертности в случайно скрещивающейся популяции, но также и все средовые компоненты 1).
Вообще говоря, данные таблиц 6.20 и 6.21 довольно неутешительны. Из-за того что отношение В/А варьирует в широких пределах, мы даже не можем с уверенностью сказать, что инбридинг как таковой повышает риск мертворождения и детскую смертность, хотя такое утверждение и представляется вероятным. Однако один вывод очевиден. Высокие значения отношения В/А, полученные для данных, проанализированных Мортоном с соавт. [1827], не подтвердились ни в одной из других работ. Вероятно, они завышены в результате методических отклонений или из-за социально-экономических различий между кровнородственными и некровнородственными браками.
Рецессивные болезни и врожденные уродства в потомстве от кровнородственных браков. Наше обсуждение было пока совершенно абстрактным. Если дети от кровнородственных браков чаще рождаются мертвыми или гибнут в раннем возрасте, возникает вопрос: почему это происходит? Страдают ли они от известных рецессивных заболеваний или от мультифакториальных пороговых болезней, таких, как врожденные аномалии.
Наиболее полные данные по этому вопросу были получены японскими исследователями (табл. 6.23 [1876]). В этой таблице приведены частоты грубых врожденных аномалий, полученные в одной из работ японских авторов. Между кровнородственными и некровнородственными браками наблюдается незначительное достоверное различие. В той же большой когорте новорожденных Японии суммарная частота смертных случаев и грубых врожденных аномалий составила 4,3% у контрольных детей и 6,2% у детей от браков между двоюродными сибсами. Большинство аномалий в этой работе являлись уродствами (иногда очень сложной природы), рецессивное наследование которых доказано не было. Даже из наиболее подробного обзора [198] нельзя установить приблизительное число идентифицируемых заболеваний с точно установленным рецессивным типом наследования. Это означает, что в результате огромной работы по изучению влияния кровного родства не было получено данных, позволяющих приписать хотя бы некоторые из эффектов инбридинга определенной группе генов, которые в рецессивном состоянии приводят к возникновению болезней.
Другие признаки, проявляющие эффект инбридинга: умственные способности. В нескольких работах изучалось влияние кровного родства на антропологические
1) Кроме этих отклонений отношение В/А зависит также от особенностей статистической оценки [1763].
6. Популяционная генетика 357
Таблица 6.23. Зависимость числа детей с тяжелыми врожденными аномалиями от места жительства и степени кровного родства родителей [1876] |
||||||
Город |
См. сноску" |
Браки двоюродных сибсов |
Браки двоюродный дядя х племянница |
Браки троюродных сибсов |
Некровнородственные браки |
Сумма |
Хиросима |
п |
936 |
313 |
384 |
26012 |
27 645 |
|
т |
17 |
2 |
4 |
293 |
316 |
|
Р |
0,0182 |
0,0064 |
0,014 |
0,0113 |
0,01114 |
Куре |
п |
318 |
113 |
140 |
7544 |
8115 |
|
т |
4 |
2 |
1 |
58 |
565 |
|
Р |
0,0126 |
0,0177 |
0,0071 |
0,0077 |
0,0080 |
Нагасаки |
п |
1592 |
412 |
637 |
30240 |
32 881 |
|
т |
27 |
4 |
8 |
300 |
339 |
|
Р |
0,0170 |
0,0097 |
0,0126 |
0,0099 |
0,0103 |
Всего |
п |
2846 |
838 |
1161 |
63 796 |
68 641 |
|
т |
48 |
8 |
13 |
651 |
720 |
|
Р |
0,0169 |
0,0095 |
0,0112 |
0,0102 |
0,0105 |
|
|
|
Анализ 2) |
|
|
|
|
|
|
χ2 |
DF |
Р |
|
Города |
|
|
7,269 |
2 |
0,02 < Р < 0,05 |
|
Кровнородственные—некровнородст венные браки |
11,775 |
3 |
0,001 < Р < 0,01 |
|||
Взаимодействие |
|
2,535 |
6 |
0,75 < Р < 0,90 |
||
1) n число детей; т- число детей с тяжелыми врожденными аномалиями. 2) Использован метод Роя и Кастенбаума (1956); см. Шелл (1958). Между частотами основных врожденных нарушений в кровнородственных и некровнородствениых браках наблюдается значимое различие. |
||||||
признаки, стоматологические характеристики, кровяное давление, координацию, остроту зрения и слуха, умственное развитие, школьную успеваемость [1840; 1879; 198; 1765]. В целом значение каждого признака немного уменьшалось с увеличением инбридинга. Особенно интересны результаты, касающиеся умственного развития и школьной успеваемости, которые только частично обусловлены социально-экономическими различиями. После исключения путем соответствующей статистической обработки социально-экономических факторов было показано следующее: при увеличении F на 10% коэффициент интеллектуального развития IQ уменьшался на 6 единиц по вербальным субтестам и субтестам действия при использовании шкалы Векслера для детей (WISC). В такой же степени снижалась и успеваемость в школе [198].
Заключение о том, что инбридинг понижает средний уровень познавательных способностей, подтверждается исследованием выборки из 3203 школьников арабов в Израиле. В этой популяции процент браков между двоюродными сибсами составлял около 34%; приблизительно 4% браков были между дважды двоюродными сибсами. Тщательно проанализированные социально-экономические условия оказались практически идентичными в инбредной группе и неинбредном контроле. Средние результаты трех различных тестов по проверке умственных способностей оказались значительно ниже у детей от браков двоюродных сибсов (и особенно дважды двоюродных сибсов) по сравнению с
358 6. Популяционная генетика
контрольной группой. Те же различия наблюдались по средней школьной успеваемости по четырем предметам [1723].
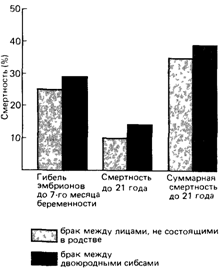
Общая оценка потери гетерозигот, обусловленная кровным родством родителей. Данные по влиянию кровного родства на умственное развитие или антропометрические признаки интересны со многих точек зрения. Однако они не увеличивают наших знаний о влиянии кровного родства родителей на гибель зигот до достижения репродуктивного возраста, а это единственный параметр, важный для определения летального эквивалента. В табл. 620 и 6.21 приведены сведения только о перинатальной смертности. Эти данные на включают гибель эмбрионов до седьмого месяца беременности, а также детей после достижения 8-летнего возраста. Последней группой можно пренебречь, так как в большинстве развитых стран смертность в ней очень низка. Относительно спонтанных абортов имеются только отрывочные сведения. Соответствующая оценка Шелла и Нила (1972) [1879] приведена на рис. 6.43.
|
Рис. 6.43. Потеря зигот в связи с кровным родством родителей в результате спонтанных абортов и ранней смертности (от 8 мес беременности до 21 г.) [1879]. |