

5.1.4.4. Мутации у микроорганизмов: их вклад в понимание механизма мутаций у человека
Мутации как ошибки репликации ДНК. Данные, полученные на человеке, свидетельствуют о существовании тесной связи между мутациями и клеточными делениями. Импульсом к изучению этой проблемы послужила гипотеза о механизме возникновения точковых мутаций Уотсона и Крика (рис. 5.26) [1347]. Важную роль в этом сыграли и ранние исследования на микроорганизмах, из которых следовало, что многие спонтанные мутации действительно возникают во время репликации ДНК в результате ошибочной вставки неправильного нуклеотида, приводящей к появлению в будущих клеточных поколениях новой, отличающейся пары оснований. В последние десятилетия получено до удивления мало новых данных о механизмах возникновения спонтанных мутаций, что очень сильно контрастирует с громадным объемом имеющихся в настоящее
192 5. Мутации
|
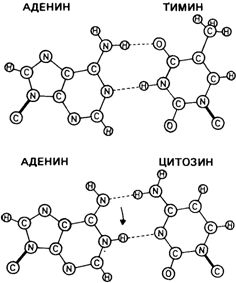
Рис. 5.26. Механизм возникновения точковой мутации в результате замены основания (в соответствие с моделью Уотсона -Крика). Предполагается, что любое основание на короткое время может принимать редкую таутомерную конфигурацию и образовывать пару не со своим обычным партнером, а с другим основанием, например аденин может спариться не с тимином, а с цитозином. Ко времени следующего репликационного цикла оба основания примут свои наиболее вероятные конфигурации и образуют пары со своими обычными партнерами. Следовательно, две молекулы следующего поколения будут отличаться от своих предшественников, т. е. произошла точковая мутация [1347]. |
время материалов о механизмах индуцированного мутагенеза.
Учитывая важность спонтанных мутаций для эволюции (разд. 7.2.3) и для любых прогнозов биологического будущего человеческого вида, трудно понять, почему этих данных так мало. Наиболее очевидное объяснение состоит в том, что эволюционисты-теоретики обычно не принимают участия в планировании экспериментов по генетике микроорганизмов. Кроме того, изучение спонтанных мутаций в большинстве случаев отнимает намного больше времени, чем эксперименты по индуцированному мутагенезу. Тем не менее коекакими данными такого рода мы располагаем. Как и предполагалось, аминокислотные замены, возникшие в результате спонтанной точковой мутации в локусе триптофансинтетазы А Е. coli, обусловлены замещением одного основания точно так же, как и точковые мутации гемоглобина [1703].
Детальному анализу подверглись мутации в области rII бактериофага Т4 [53]. Преобладающее большинство из них связано с репликацией и представляет собой сдвиги рамок считывания. Особенно часто такие мутации возникают в двух горячих точках. Если исключить из рассмотрения мутации, происходящие в них, отношение частот сдвигов рамок считывания к одиночным нуклеотидным заменам снижается с 3,3 до 1,6. Данные, свидетельствующие о том, что и у бактерий мутации зависят от репликации, представлены в работе Кондо [1524].
Таким образом, у микроорганизмов многие мутации, вероятно даже подавляющее большинство, связаны с репликацией. Часть из них вызвана заменой одного основания, в других случаях мутации – результат сдвига рамки считывания. Это означает, что объяснение эффекта отцовского возраста и различий по полу (разд. 5.1.3) ошибочным спариванием оснований в процессе репликации хотя и может быть правильным во многих случаях, не обязательно должно касаться всех мутаций.
В течение долгого времени возможность возникновения каких-либо репликационнонезависимых спонтанных мутаций казалась сомнительной. Было известно о накоплении мутаций в неделящихся гаметах Drosophila, в спорах Neurospora и у E. coli в стационарной фазе. Однако в этих случаях мог иметь место репаративный синтез ДНК. Определенный ответ в конце концов был получен в исследованиях на бактериофаге Т4 [53]. В свободных фаговых частицах не происходит ни репарации, ни репликации, однако было обнаружено, что в определенных rII-мутантах с течением времени имеет место линейное накопление реверсий. Однако их частота была низкой по сравнению с частотой при репликации. В случае репликационно-независимых мутаций сдвиги рамок считывания были редки, а трансверсии происходили чаще, чем в случае репликационно-зависимых мутаций.
5 Мутации 193
Гены-мутаторы. В 1937 г. Демерец [1435] описал «нестабильные» гены в определенных линиях Drosophila melanogaster. С тех пор стали известны многочисленные примеры генетически детерминированных, необычайно высоких мутационных частот как у эукариот, так и у прокариот. Причины этой повышенной мутабильности часто можно связать с «геном-мутатором». Анализ влияния таких генов дал ценную информацию о взаимодействии различных факторов (полимераз, ферментов репарации и т.д) [53; 1439; 1558], участвующих в мутационном процессе. В случае точковых мутаций в половых клетках нет никаких данных, свидетельствующих о том, что такие гены-мутаторы действительно существуют. Было бы интересно провести тщательный поиск крайне редких человеческих семей с двумя мутациями. Однако при анализе мутаций в отдельных клетках гены-мутаторы были идентифицированы (см. ниже).
Происходят ли мутационно-подобные события под влиянием вирусов и транспозонов? До сих пор спонтанные мутации рассматривались в свете классических концепций (замена основания, делеция, рекомбинация). Однако при обсуждении новых результатов, касающихся структуры хромосом и ДНК у человеческих индивидов (разд. 2.3), мы упоминали также о транспозонах или «прыгающих генах». Фенотипически их проявления неотличимы от эффектов классических мутаций. Мы не знаем, играют ли транспозоны какую-либо роль при возникновении и передаче мутаций в половых клетках человека. Однако, поскольку они обнаружены не только у прокариот [520], но также и у некоторых эукариот (см. разд. 2.3.4), их присутствие у человека представляется вполне вероятным.
Латентные вирусы могут передаваться вертикально от поколения к поколению, не оказывая какого-либо вредного действия. Вместе с тем они могут влиять на физиологию своих хозяев Все клубни картофеля сорта Кинг Эдуард несут вирус морщинистой курчавости листьев, не проявляя при этом симптомов заболевания, однако растения, не содержащие вируса, отличаются по внешнему виду от растений обычных линий и дают более высокий урожай. В системах in vitro было показано, что некоторые патогенные вирусы, например вирус немецкой кори, индуцируют хромосомные аберрации у человека [1386, 1479] Такие аберрации можно также индуцировать в хромосомах клеток зародышевого пути (в 1-м мейотическом делении) самца мыши [1655]. Еще в 1963 г. Тейлор [1654] обнаружил, что фаг Mu E. coli индуцирует множество генных мутаций в различных сайтах. Впоследствии этот фаг был назван транспозоноподобным. Как выяснилось, многие спонтанные фаговые мутации имеют транспозонную природу. В последующем было обнаружено, что вирусы животных, например SV40 и вирус полиомы, способны индуцировать генные мутации в клетках млекопитающих (в клеточных линиях китайских хомячков и мышей) [1463]. Как уже отмечалось (разд. 5.1.3.3), частоты многих доминантных мутаций заметно растут с увеличением возраста отцов. Но в некоторых случаях эффект отцовского возраста настолько слабый, что невозможно решить, обусловлен ли он действительно возрастом отцов, матерей или и тех и других. Соответствующий пример приведен на рис. 5.19, где показано увеличение частоты ретинобластомы. К доминантным заболеваниям, проявившим лишь небольшое увеличение частоты, относятся нейрофиброматоз, туберозный склероз и полипоз кишечника. Все эти заболевания - опухолевые. Вирусная индукция опухолей в соматических тканях и роль онкогенов будут рассматриваться в разд. 5.1.6 5. Здесь выскажем только предположение, что в случае перечисленных выше болезней именно мобильные элементы или вирусы могут вызывать в клетках мутационноподобные события.