

5.1.3.5. Герминативноклеточные и соматоклеточные мозаики по доминантным и х-сцепленным мутациям
Анализ родословных. Если мутация происходит во время раннего развития половых клеток, возможно образование герминативных мозаиков, у которых более или менее значительная часть клеток одной из гонад несет эту мутацию. Такая ситуация хорошо известна из работ по изучению мутагенеза у Drosophila melanogaster; кроме того, у мыши описан «кластер» клеток, возникший в результате мутации на ранних стадиях развития ооцита (см. табл. 5.15). Вероятность обнаружения таких кластеров у людей очень мала; их можно выявить только в том случае, если мутация имеется в клетках, составляющих большую часть гонады.
|
Рис. 5.21. Зависимость числа мутантных половых клеток от стадии развития, во время которой произошла мутация. |
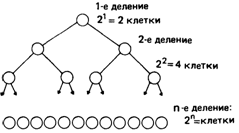
Чем раньше в процессе развития возникает мутация, тем больше мутантная часть гонады. Доля мутантных половых клеток равна 1, если уже первая клетка оказывается носителем мутации; она составляет 1/2, если мутация произошла при первом делении стволовой клетки, и (1/2)2, если мутация возникла во время второго деления; в общем случае, если мутация произошла в n-м делении, она равна (1/2)n (рис. 5.21). Если общее число клеточных делений по порядку величины совпадает с полученной выше оценкой (разд. 5.1.3.3; рис. 5.15 и 5.16), а вероятность мутации одинакова для всех клеточных делений, то доля мутаций de novo, образующих кластеры, выявляемые при герминативном мозаицизме, будет хотя и небольшой, но все же ощутимой. Если мутация доминантна, можно ожидать появления немногочисленных семей, в которых оба родителя нормальны и в то же время имеется более одного ребенка с мутантным фенотипом.
Описано несколько редких родословных, в которых, вероятно, возникли такие кластеры, в частности одна большая родословная с аниридией [1596] и семья, где есть больные с расщепленной кистью и стопой [1542]. Во всех подобных случаях очень трудно исключить более тривиальную альтернативу, а именно неполную пенетрантность у одного из родителей.
Соматическая мозаичностъ в случае доминантных мутаций. Мозаичность, возникшая в результате мутации, бывает не только в герминативной, но и в соматической ткани. Последняя может быть мозаична не
184 5. Мутации
только по числу хромосом, о чем говорится в разд. 5.1.6, но и по генным мутациям. Однако характер фенотипического проявления генных мутаций сильно затрудняет обнаружение такого мозаицизма. Тем не менее по крайней мере один такой факт нам известен. В ходе изучения нейрофиброматоза [1426] обнаружено четыре индивида, имеющих нейрофиброму, ограниченную какой-либо одной частью тела, например конечностями, крестцовым отделом или спиной. В семейных историях этих четырех индивидов подобных больных не было. Они произвели в общей сложности шесть детей, ни один из которых, по имеющимся сведениям, не страдал данной болезнью. Следовательно, у этих четырех индивидов возникли соматические мутации, оказавшие влияние на относительно ранние стадии развития.
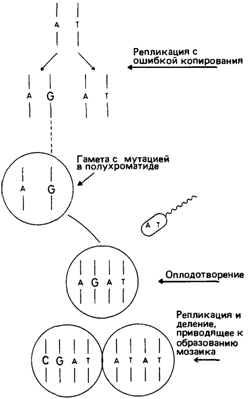
Мутации в полухроматидах? Недавно Гартлер и Франке (1975) [676] выдвинули гипотезу о существовании специального механизма образования мозаиков по точковым мутациям, связанного с мутациями в полухроматидах (рис. 5.22). Как отмечалось выше, многие мутации, по-видимому, возникли вследствие ошибок копирования при репликации ДНК. Если такая ошибка копирования происходит в последнем перед образованием половой клетки цикле репликации ДНК, возникшая половая клетка будет содержать неспаренную пару оснований, например AG вместо AT (рис. 5.22). В первом делении дробления А образует пару с Т, a G – c С. Следовательно, один из продуктов этого деления будет, как и прежде, содержать пару оснований AT; другая же клетка получит новую пару оснований GC и будет мутантной.
Примером таких мозаиков могут служить больные пигментным дерматозом. Как мы уже говорили, это заболевание, вероятно, обусловлено доминантным геном, сцепленным с Х-хромосомой и летальным у гемизиготных мужчин. Всего было описано 593 случая этой болезни у женщин и шесть у мужчин, имевших нормальные кариотипы XY. Распределение пораженных участков кожи было сходным у индивидов обоего пола и напоминало мо-
|
Рис. 5.22. Возникновение мутации в полухроматиде: замена основания в одной полухроматиде, происходящая во время последнего перед мейозом цикла репликации ДНК. В первом делении дробления мутантная полухроматида дает начало клону клеток, имеющих генную мутацию; нормальная полухроматида приводит к образованию клона нормальных клеток. Данный индивид будет мозаиком с соотношением нормальной и мутантной ткани 1:1. |
заичный рисунок на шкурках мышей, хомячков и кошек, гетерозиготных по некоторым Х-сцепленным генам.
Кроме того, случаи заболевания мужчин носили спорадический характер. Тот факт, что больные мужчины обнаруживают большое фенотипическое сходство с женщинами-мозаиками, мозаицизм которых обусловлен эффектом Лайон (см. разд. 2.2.3.3), делает вероятным предположение, что эти мужчины также являются мозаиками и что соответствующая мутация произошла на ранней стадии эмбрионального
5. Мутации 185
развития. Подобные случаи хорошо объясняются гипотезой о мутациях в полухроматидах [760].