

4.3.5. Популяциоииая генетика генов гемоглобина (см. [972], разд. 6.1.2.3)
Обнаруженный в локусе Hbβ полиморфизм позволяет проследить возникновение и распространение различных гемоглобинопатии. Можно предположить, что мутация «гемоглобин S» возникла независимо в трех различных географических зонах (Сенегал, Бенин, Банту) и затем широко распространилась благодаря селективному преимуществу, которое она дает против заболевания малярией [1833]. Среди хромосом с мутацией НbЕ в Юго-Восточной Азии различают три гаплотипа, один из которых возникает в результате кроссинговера в «горячей точке» (см. выше). Мутация «гемоглобин С» сцеплена, как правило, с одним гаплотипом (см., однако, разд. 6.2.1.6). Отдельные мутации, приводящие к β-талассемии, обычно возникают в уникальных гаплотипах и затем распространяются благодаря селекции на устойчивость к малярии. Из 31 точковой мутации, приводящей к β-талассемии, лишь 2 встречаются более чем у одной этнической группы. Связь конкретной мутации со специфическим гаплотипом в значительной степени облегчила установление природы различных мутаций, вызывающих талассемию. Поскольку для исследования были выбраны различные гаплотипы ДНК, существовала вероятность обнаружения новых, ранее неизвестных мутаций. И действительно, удалось открыть много новых типов талассемии [1253]. Можно заключить, что все распространенные талассемии и гемоглобинопатии возникли уже после дивергенции человеческих рас и в большинстве случаев остаются сцепленными с хромосомами, в которых они впервые появились. Если одна и та же мутация сцеплена с разными гаплотипами, это в большинстве случаев проще всего объяснить мейотическим кроссинговером в «горячей точке» рекомбинации, расположенной перед геном β-глобина.
Большой интерес вызывает возможность межаллельной конверсии. Так, мутация Нbβ39 на острове Сардиния сцеплена с несколькими гаплотипами, что трудно объяснить простой рекомбинацией. Более вероятным механизмом считается однонаправленный перенос генетической информации, или конверсия (рис. 2.97). Этот же механизм предполагают для объяснения подобных данных по мутациям гемоглобин S, гемоглобин Е и мутации сдвига рамки в гене β-глобина. Такой механизм представляется очень привлекательным, поскольку имеются убедительные доказательства генной конверсии в эволюции генов глобинов у нечеловекообразных обезьян. Высказываются предположения о молекулярных механизмах такой конверсии [972]. Для шести хорошо изученных мутаций в интронах, которые приводят к талассемии, признаков генной конверсии не обнаружено.
4.3.6. Пренатальная диагностика гемоглобинопатии [966; 2269; 2322; 2361]
Существует несколько возможных подходов к пренатальной диагностике гемо-
4. Действие генов 99
|
|
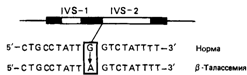
Рис. 4.59, Олигонуклеотидный зонд (олигонуклеотид из 19 остатков) для гена нормального β-глобина отличается от зонда для гена с мутацией, приводящей к β -талассемии только заменой G А в интроне IVS-2. В соответствующих условиях гибридизации зонд для мутантного гена будет узнавать только мутантный ген, но не нормальный. Аналогичным образом «нормальный зонд» не будет гибридизоваться с мутантным геном. |
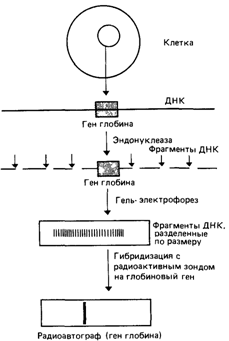
Рис. 4.60. Обнаружение глобинового гена. ДНК выделяют из любого препарата клеточных ядер, в ее составе имеются глобиновые гены. Различные рестрикционные эндонуклеазы (рестриктазы) расщепляют ДНК на множество фрагментов (рестриктов), узнавая специфические последовательности нуклеотидов. Фрагменты разделяют по размеру с помощью гель-электрофореза. Готовят специфический радиоактивный зонд на глобиновый ген и гибридизуют его с фрагментами ДНК Радиоактивный сигнал обнаруживают с помощью радиоавтографии [1252]. |
глобинопатий: а) биохимический анализ гемоглобина на уровне белка в образцах крови, получаемых при фетоскопии или пункции плаценты; б) амниоцентез (пункция амниотической жидкости) на 15-16-й неделе с последующим анализом ДНК амниотических клеток; в) анализ ДНК ворсинок хориона, полученных при биопсии хориона на 9-10-й неделе беременности (разд. 9.1.1). В прямой диагностике клеток и тканей плода используются рестрикционные эндонуклеазы (рестриктазы), позволяющие распознать специфические мутации, например гемоглобин S, а также олигонуклеотидные зонды [1254; 2376], способные гибридизоваться с ДНК при конкретной мутации, вызывающей талассемию (рис. 4.59). В основе косвенных методов пренатальной диагностики лежит семейный анализ рестрикционных сайтов ДНК (выявляемых при анализе полиморфизма длины рестрикционных фрагментов), тесно сцепленных с изучаемой формой талассемии (разд. 9.1.1).
Все эти методы были использованы к настоящему моменту. Взятие образцов крови при фетоскопии или пункции плаценты во многих случаях приводит к гибели плода (5% у опытных специалистов), хотя сам анализ относительно прост: для крови, взятой непосредственно у плода, достаточно методов исследования белков. По мере усовершенствования и упрощения методов молекулярной биологии (рис. 4.60) все большее число лабораторий переключается на их использование. Биопсия хориона имеет некоторые преимущества перед амниоцентезом: при этом удается получить больше ДНК, на более ранних сроках, меньше времени требуется для окончательного заключения. После амниоцентеза для получения биомассы, необходимой для анализа ДНК, клетки приходится культивировать в течение нескольких недель.
Методы прямого анализа ДНК всегда более предпочтительны, поскольку не связаны с изучением семей. Так, в диагностике талассемии большие надежды возлагают на применение специфических олигонуклеотидных зондов и разработку методов гибридизации без использования радиоактивной метки. Однако при этом заранее необходимо предполагать, какой именно му-
100 4. Действие генов
тации, приводящей к талассемии, следует ожидать. В этом могут помочь данные по сцеплению конкретной мутации со специфическим гаплотипом ДНК. Таким образом, для установления природы мутации, вызывающей талассемию, требуется предварительное исследование ДНК больного родственника.
Гемоглобин как модельная система. Гемоглобин - наиболее изученная генетическая система у человека. На основе концепций, разработанных в ходе ее изучения, можно глубже понять другие явления в генетике человека. Например, если в разных семьях обнаруживаются наследственные заболевания с различным фенотипическим проявлением, обычно заключают, что они вызваны мутациями в разных генах. Исследования гемоглобина показывают, что так бывает не всегда. Например, хотя метгемоглобинемия фенотипически сильно отличается от гемолитической анемии или эритроцитоза, причиной их являются аллельные мутации. Таким образом, фенотип определяется тем, какая именно молекулярная аномалия лежит в его основе и каким образом при этом изменена нормальная функция.
Другой полезный урок можно извлечь из того, как тетрамерная структура гемоглобина обеспечивает функцию связывания кислорода, а также как мутации могут влиять на эту функцию, нарушая взаимодействие различных глобиновых цепей. Хотя большинство мутаций, изменяющих гемоглобин, нейтральны, все патологические варианты, за исключением серповидноклеточной анемии, наследуются по доминантному типу. Отсюда следует, что один из возможных механизмов доминирования заключается в нарушении взаимодействия между продуктами аллельных генов (разд. 4.6).
Наконец, изучение гемоглобинов продемонстрировало многообразие механизмов возникновения мутаций у человека. Они могут затрагивать как структурные гены, так и прилегающие регуляторные участки. В большинстве случаев - это замены нуклеотидов, но встречаются и делеции, которые могут сильно различаться по длине. Хотя у прокариот мутации со сдвигом рамки считывания широко распространены, их обнаружение у человека было несколько неожиданным для специалистов. Наше представление о роли мутаций в эволюции во многом основано на результатах изучения мутаций гемоглобина.