

4.3. Гемоглобин человека [119; 31; 97а]
Молекулу гемоглобина изучать легче, чем молекулу любого другого белка человека. Гемоглобин – основной белок эритроцитов, и для его выделения не требуется сложных биохимических методов. Неудивительно поэтому, что именно об этом белке мы знаем больше, чем обо всех остальных. Исследования по генетике гемоглобина человека, изучение аминокислотной последовательности и структуры его молекулы продвигались очень быстро. В молекулярной генетике человека они сыграли такую же роль, как изучение дрозофилы и бактериофагов в общей генетике. Большинство концепций, разработанных для этой системы, являются общими для других белков. Действительно, многие концептуальные принципы генетики человека можно иллюстрировать примерами из генетики гемоглобина.
4.3.1. История изучения гемоглобина
Серповидноклеточная анемия – «молекулярное» заболевание. Изучение гемоглобина человека началось с открытия наследственного заболевания – серповидноклеточной анемии. В 1910 г. Херрик [1121] обнаружил у студента-негра, страдающего анемией, особую аномалию эритроцитов: они были серповидной формы. Вскоре выяснилось, что такая патология довольно часто встречается у американских негров. Больные страдали от гемолитической анемии и частых болей в кишечнике и скелетных мышцах. Было показано, что больные серповидноклеточной анемией гомозиготны по гену, который в гетерозиготном состоянии (примерно у 8% американских негров) вызывает гораздо менее выраженное отклонение: присутствие в крови некоторого количества серповидных эритроцитов [1226].
Решающую роль в биохимическом и генетическом анализе этой болезни сыграла работа выдающегося химика Полинга,
4 Действие генов 71
|
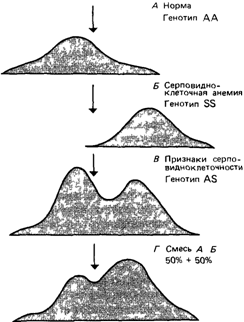
Рис. 4.33. Диаграмма зонального электрофореза гемоглобинов при рН = 6,9. А Нормальная гомозигота (АА) Б Больной с серповидноклеточной анемией (SS) В Признак серповидноклеточности (AS) Г Смесь равных количеств гемоглобина А и гемоглобина S [1260] Стрелка указывает на стартовую точку электрофореза |
опубликованная под программным заголовком «Серповидноклеточная анемия, молекулярное заболевание» [1260] (Полинг узнал об этой болезни от Кастла, известного гематолога и сына одного из пионеров генетики млекопитающих, и предположил, что ее причиной может быть дефект гемоглобина ) Он писал
«Данные, имевшиеся к началу нашей работы, указывали, что процесс образования серповидных эритроцитов может быть тесно связан с состоянием и природой гемоглобина в эритроцитах»
Авторы исследовали гемоглобин людей, в крови которых обнаруживались серповидные эритроциты, гемоглобин больных серповидноклеточной анемией и гемоглобин здоровых людей В работе использовали самый совершенный в то время метод анализа белков – зональный электрофорез по Тизелиусу (рис 4 33) Пики на рисунке соответствуют градиентам концентрации гемоглобина в определенном буфере, расположение этих пиков зависит от соотношения положительных и отрицательных зарядов в молекуле белка
«Результаты указывают на существование значительных различий в электрофоретической подвижности гемоглобина, выделенного из эритроцитов здоровых людей, и гемоглобина, выделенного из эритроцитов больных серповидноклеточной анемией»
У людей, в крови которых наряду с нормальными имеются и серповидные эритроциты, обнаружено 25-40% аномального гемоглобина, такого же как у больных серповидноклеточной анемией, остальной гемоглобин был неотличим от гемоглобина нормальных индивидов Эти данные подтверждали предположение о том, что больные серповидноклеточной анемией гомозиготны по гену, который находится в гетерозиготном состоянии у людей с признаком серповидноклеточности
«Эта работа показала, что молекула белка меняется при аллельном изменении единственного гена, контролирующего его синтез»
Замена одной аминокислоты В 1956 г Ингрэм работал в Кэмбридже, в той лаборатории, где до этого Перутц исследовал кристаллографию белков, Сэнгер определил аминокислотную последовательность инсулина, а Крик и Уотсон предложили свою модель структуры ДНК Ингрэму удалось точно определить, чем нормальный гемоглобин отличается от серповидноклеточного [1138] При гидролизе молекулы глобина трипсином, расщепляющим белки, образуется около 60 пептидов, которые были разделены в двумерной системе на бумаге в одном направлении с помощью электрофореза, а в другом – с помощью хроматографии Этим методом (его называют методом «отпечатков пальцев») удалось показать, что гемоглобин серповидных эритроцитов отличается от нормального по подвижности единственного пептида При дальнейшем анализе этого пептида выяснилось, что гемоглобин серповидных эритроцитов отличается от нормального только по одной аминокислоте, глутамино-
72 4. Действие генов
вая кислота в определенном положении заменена валином.
|
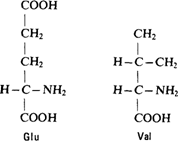
В молекуле глутаминовой кислоты по сравнению с молекулой валина имеется дополнительная карбоксильная группа. Эта разница в зарядах и обусловливает различия в электрофоретической подвижности нормального и серповидноклеточного гемоглобина.
Впоследствии, по мере совершенствования методов электрофореза, стали выявляться все новые и новые варианты гемоглобина. В настоящее время их известно более 400 [1194]. Следующими вехами в изучении гемоглобина следует считать установление его полной аминокислотной последовательности (Браунитцер и др., 1961) [1016] и трехмерной структуры [1165; 1265]. Позже стали понятны структурнофункциональные взаимоотношения, были обнаружены различные типы мутаций: делеции и сдвиг рамки считывания. Выделение мРНК гемоглобина позволило по-новому взглянуть на структуру и функционирование гена, открыло новые пути к пониманию механизма его действия.
Исследования гемоглобинов на молекулярном уровне продвигались очень быстро. В настоящее время известны полные нуклеотидные последовательности ряда генов гемоглобинов вместе с фланкирующими их последовательностями, мы хорошо понимаем организацию гемоглобиновых генов, изучена природа мутаций, затрагивающих гемоглобины, в особенности при талассемиях. Следующий раздел посвящен генетике гемоглобинов.