

Ряд Морські ковпаки (Thalassocolycida)
Це дуже ніжні істоти, що нещодавно були знайдені в Саргасовому морі. Своїм куполоподібним тілом вони нагадують медуз; живляться пасивно, збираючи ротом планктонні організми, що прилипають до внутрішньої поверхні купола. Рот міститься на особливому вирості, й до нього підтягуються різні ділянки купола.
Реброплави за рівнем організації близькі до кишковопорожнинних і раніше об'єднувалися з ними в один тип. Але вони істотно відрізняються від кишковопорожнинних тим, що мають аборальний орган, рухаються переважно за допомогою війчастих утворень, не мають прикріпленої стадії в життєвому циклі, замість жалких мають клейкі клітини. Реброплави — єдина група багатоклітинних, яка зберегла первиннопланктонний спосіб життя й походить від вільноплаваючих предків. Кишковопорожнинні, навпаки, перейшли до прикріпленого способу життя, втратили аборальний орган і здатність до війчастого руху, що залишилася тільки в личинок. Отже, реброплави та кишковопорожнинні хоча й мають спільних предків, розвивалися різними еволюційними шляхами.
Реброплави заслуговують на особливу увагу дослідників у зв'язку з тим, що принцип їх будови часто використовується для вирішення важливих еволюційних питань. Наприкінці минулого століття швейцарський зоолог А. Ланг обгрунтував ктенофорну гіпотезу походження плоских червів, яка базувалася на подібності будови турбелярій і повзаючих реброплавів. На його думку, сучасні війчасті черви походять від найдавніших ктенофороподібних тварин. У наш час ця гіпотеза має лише історичне значення. Вітчизняний дослідник І. І. Мечников звернув увагу на риси подібності реброплавів, личинки напівхор-дових — торнарії та личинки голкошкірих — диплеврули й висловив думку про походження вторинноротих (голкошкірих, напівхордових, хордових) від найдавніших ктенофороподібних предків.
ТИП ПЛОСКІ ЧЕРВИ (PLATHELMINTHES)
Переважна більшість плоских червів веде паразитичний спосіб життя. Вільноживучі черви трапляються в морських і пріслих водоймах і значно рідше на суходолі, у вологому грунті. Розміри плоских червів значно варіюють. Дрібні види досягають 0,5 мм, але відомі й тридцятиметрові паразити кишечника китів. Відомо близько 12 тис. видів плоских червів.
Плоскі черви — білатерально-симетричні тварини з чітко визначеним головним кінцем. У них розрізняють черевну (вентральну) та спинну (дофальну) сторони. їх тіло, як правило, сплющене в дорзовентральному напрямку, що відображає назва типу.
З
Рис. 127. Схема
поперечного розрізу плоского черва:
І—ектодерма;
2— ентодерма (середня кишка); 3
—паренхіма; 4
— кільцеві
м'язи.}
5
—
діагональні м'язи; 6
—
поздовжні м'язи; 7 — дорзо-вентральні
м'язи;
8 —
гонада;
9
—
нервовий стовбур; 10
—
статевий отвір; //— видільна пора;
12 —
протонефридій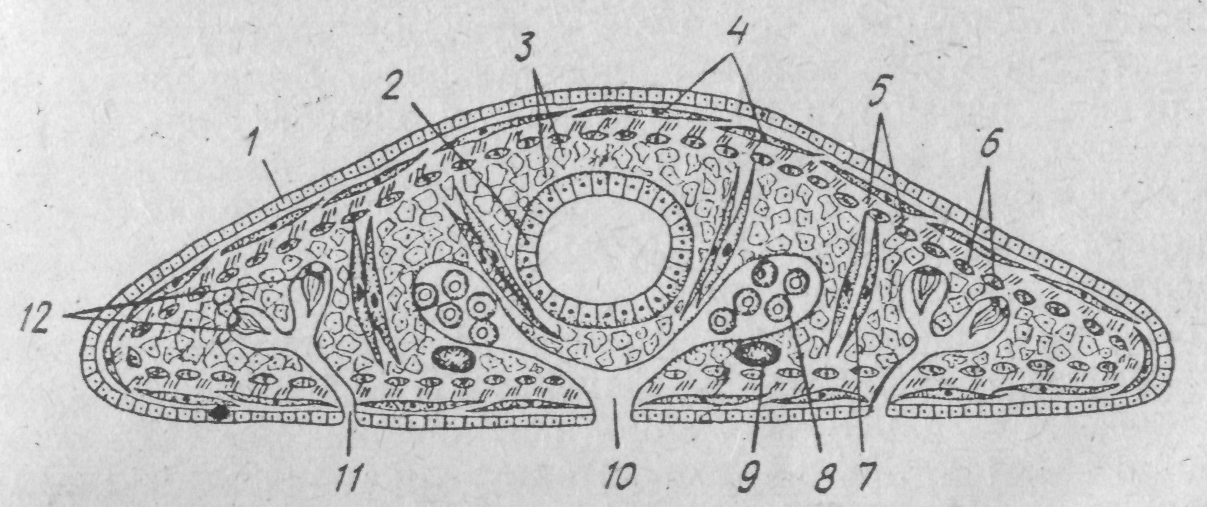 а
рівнем організації плоскі черви стоять
дещо вище кишковопорожнинних і
реброплавів. У процесі ембріонального
розвитку в них закладаються не два, а
три зародкових листки — екто-, енто-
та мезодерма. За рахунок
мезодерми формуються
мускулатура, статева система, а
також недиференційована
сполучна тканина — паренхіма
(мезенхіма), що
заповнює всі проміжки між внутрішніми
органами.
а
рівнем організації плоскі черви стоять
дещо вище кишковопорожнинних і
реброплавів. У процесі ембріонального
розвитку в них закладаються не два, а
три зародкових листки — екто-, енто-
та мезодерма. За рахунок
мезодерми формуються
мускулатура, статева система, а
також недиференційована
сполучна тканина — паренхіма
(мезенхіма), що
заповнює всі проміжки між внутрішніми
органами.
Характерною ознакою плоских червів є наявність у них шкірно-м'язового мішка. Він складається з одношарового шкірного епітелію, що має різну будову в представників різних класів, і кількох шарів м'язів — кільцевих, косих або діагональних й поздовжніх. Скороченням різних груп м'язів шкірно-м'язового мішка зумовлений червоподібний рух плоских червів. Паренхіму пронизують пучки дор-зовентральних м'язів. Крім них, є ще м'язи, що пов'язані з глоткою, органами прикріплення, статевою системою тощо.
Безпосередньо під шкірно-м'язовим мішком залягає паренхіми, що заповнює проміжки між внутрішніми органами (рис. 127). Плоскі черви належать до паренхімнихтварин і не мають порожнини тіла. Паренхіма відіграє важливу роль у житті організму. її розглядають перш за все як опорну тканину. Електронно-мікроскопічні дослідження свідчать, що в ній інтенсивно розвинена міжклітинна речовина, пронизана численними фібрилами, які мають значення опорних утворів. Клітини паренхіми розташовані рихло, між ними залишаються щілиноподібні та лакунарні простори, заповнені рідиною. Завдяки такій будові паренхіма може виконувати роль посередника в передачі продуктів травлення між кишечником і внутрішніми органами. Водночас здійснюється й транспорт продуктів обміну з міжклітинної рідини до видільної (екскреторної) системи. Останнім часом це підтверджено результатами електронно-мікроскопічних досліджень. Виявилося, що клітини паренхіми мають нерухомі вирости, які проникають у шкірно-м'язовий мішок, епітелій кишечника і видільні канали. В такий спосіб здійснюється контакт між паренхімними клітинами та внутрішніми органами. Паренхіма також є місцем накопичення поживних речовин (глікогену, ліпідів тощо). Нарешті, в паренхімі є особливі рухомі клітини, здатні до фагоцитозу, одні з них можуть виконувати захисну функцію, поглинаючи бактерії, сторонні частки, інші — екскреторну, накопичуючи тверді екскрети. тже, паренхіма — це багатофункціональна тканина.
Травна система багатьох примітивних вільноживучих видів сформована не повністю, а в деяких паразитичних зовсім редукована. У більшості плоских червів вона складається з рота, ектодермальної передньої кишки — глотки та ентодермальної сліпо замкненої середньої кишки. Часто середня кишка розгалужена, її відростки пронизують усе тіло. Таким чином, цей відділ травної системи забезпечує не лише перетравлення їжі, а й її транспорт до всіх частин тіла. Існує пряма залежність між розмірами тварини та ступенем розгалуження середньої кишки.
У плоских червів уперше з'являється спеціальна видільна система протонефридіального типу. У більшості груп є два деревоподібно розгалужених протонефридіаль-них канали з двома окремими або однією загальною порою, через які ця система каналів сполучається із зовнішнім середовищем. Внутрішні кінцеві ділянки каналів закінчуються спеціальними клітинами циртоцитами, що мають також назву зірчастих, або миготливих клітин.
Спеціальних органів дихання у плоских червів немає, як немає й кровоносної системи.
Нервова система має різну будову, але у більшості представників вона ортогонального типу — від мозкового ганглію, що розташований на передньому кінці тіла, відходять поздовжні стовбури, що з'єднуються між собою кільцевими перемичками — комісурами. Кількість поздовжніх і кільцевих нервових стовбурів варіює. У більшості плоских червів нервова система розташована не під. шкірним епітелієм, а занурена вглиб тіла.
Органи чуття представлені переважно шкірними сенси-лами, до складу яких входять нервові чутливі клітини з однією або кількома війками. Сенсили сприймають механічні та хімічні подразнення. Деякі плоскі черви мають очі та статоцисти — органи рівноваги. Місцем концентрації різних рецепторів є передній кінець тіла, де розташований мозковий ганглій, який керує їх діяльністю.
Плоскі черви за незначними винятками — гермафродити. У примітивних представників цього типу (нижчих турбелярій) немає оформлених гонад, статеві клітини розкидані в паренхімі; запліднення — внутрішнє. У більш ви-сокорозвинених форм є справжні гонади — яєчники та сім'яники, протоки для виведення сперми й зрілих яєць, а також ціла низка органів, що забезпечують внутрішнє запліднення й зберігання сперми іншої особини; формування шкаралупки яєць тощо. Деталі будови статевої системи у представників різних класів дуже різноманітні.
Характерною особливістю ембріонального розвитку плоских червів є спіральне дробіння, часто вторинно змінене.
У більшості вільноживучих турбелярій розвиток прямий, з яйця виходить особина, що відрізняється від дорослих лише за розмірами та недорозвиненою статевою системою.
Деякі морські полікладіди розвиваються з метаморфозом, у них є планктонна личинка.
У паразитичних форм, як правило, життєвий цикл дуже складний, він включає кілька поколінь, що мають різну будову. При цьому одне покоління утворюється внаслідок статевого, а друге — партеногенетичного (з незаплід-нених яйцеклітин) розмноження. Таке чергування поколінь називається гетерогонією.
Поряд із статевим розмноженням у деяких груп існує нестатеве.
Тип Плоскі черви об'єднує дев'ять класів:
Тип |
Клас |
Ряд |
|
|
Acoelida |
|
|
Macrostomida |
|
Turbellaria |
Polycladida |
|
|
Tricladida |
|
|
Rhabdocoelida |
|
|
Udonellida |
|
Xenoturbellia |
|
|
Gnathostomulida |
|
|
Trematoda |
|
Plathelminthes |
Aspidogastrea |
|
|
Monogenoidea |
|
|
Gyrocotylidea |
|
|
|
Pseudophyllidea |
|
Cestoda |
Cyclophyllidea |
|
|
Caryophyllidea |
|
Amphilinoidea |
|
До останніх шести класів належать тварини, що ведуть виключно паразитичний спосіб життя.
КЛАС ВІЙЧАСТІ ЧЕРВИ (TURBELLARIA)
Війчасті черви — це переважно вільноживучі хижаки, що мешкають у воді й лише зрідка трапляються у вологому грунті. Частина турбелярій перейшла до різних симбіотичних відносин з голкошкірими, ракоподібними, сипун-кулідами, кільчастими червами, членистоногими та рибами, є й справжні паразити. Відомо близько 3 тис. видів війчастих червів.
У прісних водоймах України знайдено близько ста видів турбелярій, приблизно стільки ж відомо з Чорного та Азовського- морів, хоча морські види вивчено гірше.
Форма тіла турбелярій листоподібна, стьожкоподібна або веретеноподібна. Більшість із них не має ніяких придатків, лише в деяких розвинені щупальцеподібні вирости на головному кінці. Розміри тіла війчастих червів переважно не перевищують 1 сантиметра, великі форми мають розмір 5—6 см, а наземні представники тропічних турбелярій, наприклад Bipalium, можуть досягати навіть 60 см завдовжки. У більшості турбелярій, перш за все морських, тіло яскраво забарвлене.
Зовні тіло турбелярій вкрите одношаровим війчастим епітелієм, що й зумовило назву класу. У примітивних без-кишкових турбелярій (ряд Acoelida) війчастий епітелій не має базальної мембрани й складається переважно з епітеліально-м'язових клітин, подібних до клітин кишковопорожнинних. Замість справжніх м'язів у них розвинені міофібрили — кільцеві та поздовжні. Базальні кінці епітеліально-м'язових клітин розхристані й занурені в паренхіму. У представників ряду Udonellida, що ведуть паразитичний спосіб життя, всі епітеліальні клітини зливаються та перетворюються на суцільну цитоплазматичну пластинку, вкриту війками. Ділянки цитоплазми з ядрами глибоко занурюються під шари м'язів. Такий тип епітелію називається зануреним. У інших представників класу епітеліальні клітини циліндричні, їх базальні кінці містяться на базальній мембрані, а вільні поверхні мають війки (рис. 128).
Б
Рис.
128. Схема будови шкірно-м'язового мішка
турбелярій — з типовим війчастим
(а)
та
зануреним епітелієм (б):
І —
клітини епітелію; 2
—
рабдити; 3
—
війки; 4
—
базальна мембрана; 5
—кільцеві,
6
—
діагональні, 7 — дорзовентральні
м'язи; 8 — поздовжні м'язи; 9
—цитоплазматична
пластинка; 10
—
занурені ділянки цитоплазми з ядрами
В епітелії або під ним міститься багато шкірних залоз. Вони виробляють слиз, що змащує тіло турбелярій. За допомогою слизу турбелярії щільно прилягають до субстрату. У деяких видів слиз використовується для здобування їжі — до тонких слизових ниток, як до павутиння, прилипає здобич. Деякі види підвішуються до поверхневої плівки води на тонкій ніжці, що утворилася із слизових виділень. Однією з різновидностей шкірних залоз є рабдитні клітини, які виробляють рабдити — блискучі палички секрету, що розташовані в епітелії перпендикулярно до його поверхні. Внаслідок подразнення тварини вони викидаються назовні, набрякають у воді й перетворюються на клейку масу. Вважають, що рабдити виконують захисну функцію та використовуються під час нападу на здобич, позбавляючи її можливості рухатися.
У деяких видів у епідермісі є спеціалізовані знаряддя нападу та захисту: гострі довгі голки, отруйні залози, секрет яких у вигляді слизу або голок чи ниток виконує захисну функцію.
П
Рис.
129. Будова травної системи різних рядів
турбелярій —Rhabdocoelida
(a);
Tricladida
— Dendrocoelum
lacteum
(б),
Polycladida
(e);
Phagocata
gracilis
(г):
/ —
ротовий отвір; 2
— глотка;
3
—
середня кишка ід
базальною мембраною містяться три шари
гладеньких м'язів (кільцеві, поздовжні
та косі, або діагональні). Сукупність
цих м'язів разом із епітелієм утворює
шкірно-м'язовий мішок. Крім того, у всіх
турбелярій є характерні для них
спинно-черевні, або дорзовентральні
м'язи. їх пучки прикріплюються до
базальної мембрани на спинній та черевній
сторонах. Таким чином тварина може
сплющувати тіло в дорзо-вентральному
напрямку.
ід
базальною мембраною містяться три шари
гладеньких м'язів (кільцеві, поздовжні
та косі, або діагональні). Сукупність
цих м'язів разом із епітелієм утворює
шкірно-м'язовий мішок. Крім того, у всіх
турбелярій є характерні для них
спинно-черевні, або дорзовентральні
м'язи. їх пучки прикріплюються до
базальної мембрани на спинній та черевній
сторонах. Таким чином тварина може
сплющувати тіло в дорзо-вентральному
напрямку.
Рух турбелярій може здійснюватися двома способами: завдяки биттю війок вони плавають, а за допомогою м'язів — повзають по субстрату та плавають, хвилеподібно вигинаючи тіло або його бічні краї. Крім того, тварина може скорочуватися й витягуватися, повертаючи тіло в різних напрямках, використовуючи при цьому різні шари м'язів шкірно-м'язового мішка. Часто турбелярії опираються на субстрат заднім кінцем і витягують тіло в поздовжньому напрямку, потім закріплюються на субстраті переднім кінцем тіла й скорочуються. Такому руху сприяє скупчення залоз з клейким секретом на передньому та задньому кінцях тіла.
Будова травної системи у представників різних рядів неоднакова (рис. 129). У безкишкових турбелярій травної системи немає. Травлення в них відбувається в паренхімі центральної частини тіла. Під ротовим отвором є скупчення спеціальних клітин — фагоцитів, у яких відбувається травлення. У більшості турбелярій є добре розвинена травна система. Починається вона ротовим отвором, що міститься на черевній стороні. Ротовий отвір веде в глотку. Іноді глоток буває дві або більше (див. рис. 129). Вони відкриваються в ентодермальний кишечник різної будови. У найбільших за розміром багатогіллястих турбелярій глотка впадає у так званий шлунок, від якого в усі боки відходять розгалужені канали, сліпо замкнені на кінцях. У тригіллястих турбелярій від глотки відходять три розгалужені гілки кишечника, одна з яких йде вперед, а дві інші — по боках глотки — назад.
У багатьох турбелярій середня кишка має вигляд мішка, а в найдрібніших прямокишкових — прямої, замкненої на кінці трубки. У турбелярій, як і в інших плоских червів, ступінь розгалуженості кишечника значною мірою залежить від розмірів тіла. Цей зв'язок пояснюється тим, що кишечник із його гілками, крім травлення, виконує функцію транспорту поживних речовин у віддалені ділянки тіла. Після надходження їжі у середню кишку паралельно з її позаклітинним травленням починається фагоцитоз часток їжі клітинами кишкового епітелію. Непере-травлені частки їжі виштовхуються в порожнину кишечника, де з'єднуються із залишками позаклітинного травлення й разом з ними викидаються назовні через ротовий отвір. При цьому турбелярія поглинає воду через рот, проганяє її через усі закутки травної системи, а потім швидким скороченням м'язів стінки тіла вимиває залишки їжі назовні.
Серед турбелярій є багато хижаків, причому вони нападають на досить велику здобич — гідр, різних червів, зокрема турбелярій інших видів, личинок комах, рачків. Часто турбелярія не проковтує жертву, а довгою глоткою проникає під покриви жертви. В її тканини виділяється спеціальний секрет глоткових залоз, що їх руйнує, після чого їжа висмоктується. Від жертви залишається тільки порожня оболонка.
Окремі види турбелярій живляться одноклітинними діатомовими водоростями, а в деяких видів у паренхімі є одноклітинні симбіотичні водорості, які синтезують необхідні турбеляріям поживні речовини.
У
Рис.
130. Протонефридіальна система — схема
загальної будови (а),
ділянка
видільної системи з циртоцитами (б),
циртоцит
(в):
/ —
видільні канальці; 2—
окремі циртоцити; З
—
видільні пори; 4
—
ядро епітеліальної
клітини; 5 — зона адсорбції
(цитоплазматичні
тяжі);
6
—
миготливе полум'я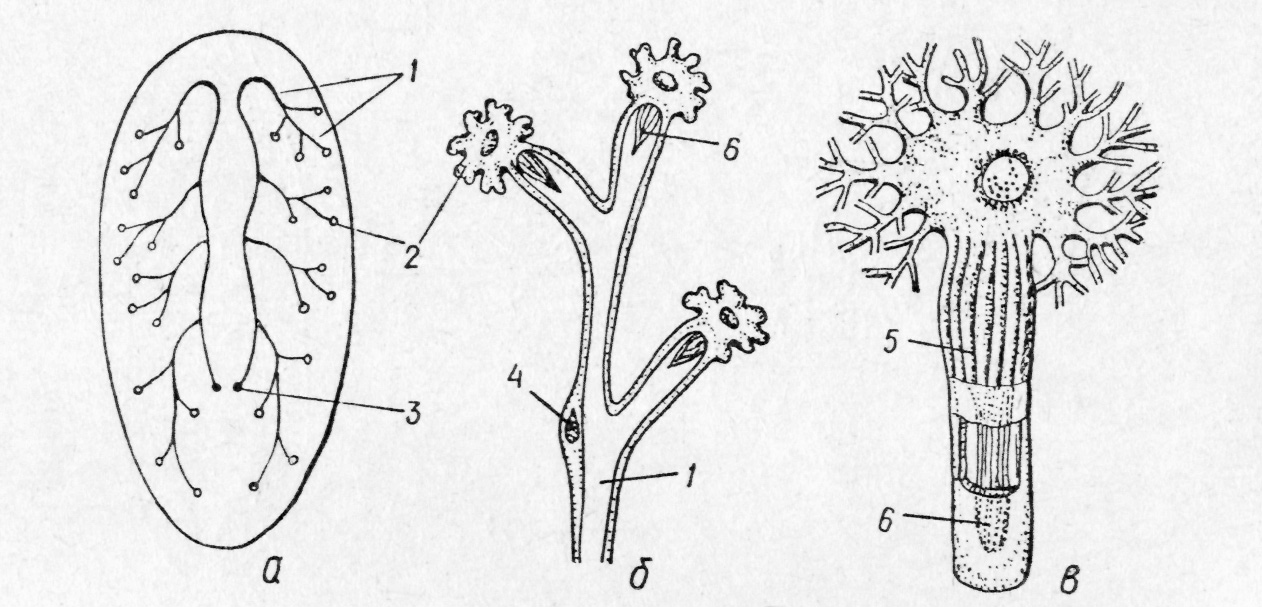 всіх турбелярій, крім безкишкових, є
типова прото-нефридіальна
видільна система (рис.
130). Вона складається з розгалужених
канальців, що починаються в паренхімі
тоненькими сліпо замкненими капілярами
всередині особливих
зірчастих клітин — циртоцитів.
У порожнину капіляра
вдається пучок війок, які весь час
рухаються, ніби полум'я свічки, що
коливається вітром, тому такі клітини
називаються миготливими,
а пучок війок всередині
них — миготливим
полум'ям. Капіляри
від різних миготливих клітин зливаються
в більші канальці, потім — у головні
канали (найчастіше їх два), що відкриваються
назовні порами.
всіх турбелярій, крім безкишкових, є
типова прото-нефридіальна
видільна система (рис.
130). Вона складається з розгалужених
канальців, що починаються в паренхімі
тоненькими сліпо замкненими капілярами
всередині особливих
зірчастих клітин — циртоцитів.
У порожнину капіляра
вдається пучок війок, які весь час
рухаються, ніби полум'я свічки, що
коливається вітром, тому такі клітини
називаються миготливими,
а пучок війок всередині
них — миготливим
полум'ям. Капіляри
від різних миготливих клітин зливаються
в більші канальці, потім — у головні
канали (найчастіше їх два), що відкриваються
назовні порами.
Електронно-мікроскопічні дослідження свідчать, що найбільш важливим у функціональному відношенні елементом протонефридіальної системи є циртоцит із капіляром. Початкова ділянка стінки капіляра має численні поздовжні щілини, затягнуті тонкою плівкою. Саме в цій ділянці міжклітинна рідина просочується до канальців. Завдяки биттю війок рідина женеться по каналах до екскреторних пор. Основна функція протонефридіїв — осморегу-ляція, тобто підтримання необхідної концентрації іонів- у міжклітинній рідині — циртоцити виводять зайву воду.
У багатьох турбелярій до цієї функції додається ще видільна. В паренхімі є спеціалізовані клітини — атроцити, в яких накопичуються кінцеві продукти обміну. Атроцити з'єднуються з каналами протонефридіальної системи, через які й виділяються накопичені екскрети і, таким чином, вона функціонує як осморегулююча та як видільна система. Поряд з цим наповнені продуктами обміну атроцити можуть також все життя лишатися в паренхімі або виво-
дитися назовні крізь стінку кишечника в його порожнину й через рот — назовні.
Н
Рис.
131. Будова нервової системи турбелярій
— схема поперечного розрізу головного
кінця турбелярії (а),
ортогон
із ендонним мозком
Convoluta (б),
ортогон
Dendrocoelum
(в):
1 —
чутливі війки, що зв'язані з нервовими
клітинами; 2—
нервові клітини;
З —
відросток нервової клітини; 4
—
мозковий ганглій; 5 — війчастий
епітелій;
6 —
статоцист; 7 — поздовжні стовбури; 8
—
поперечні нерви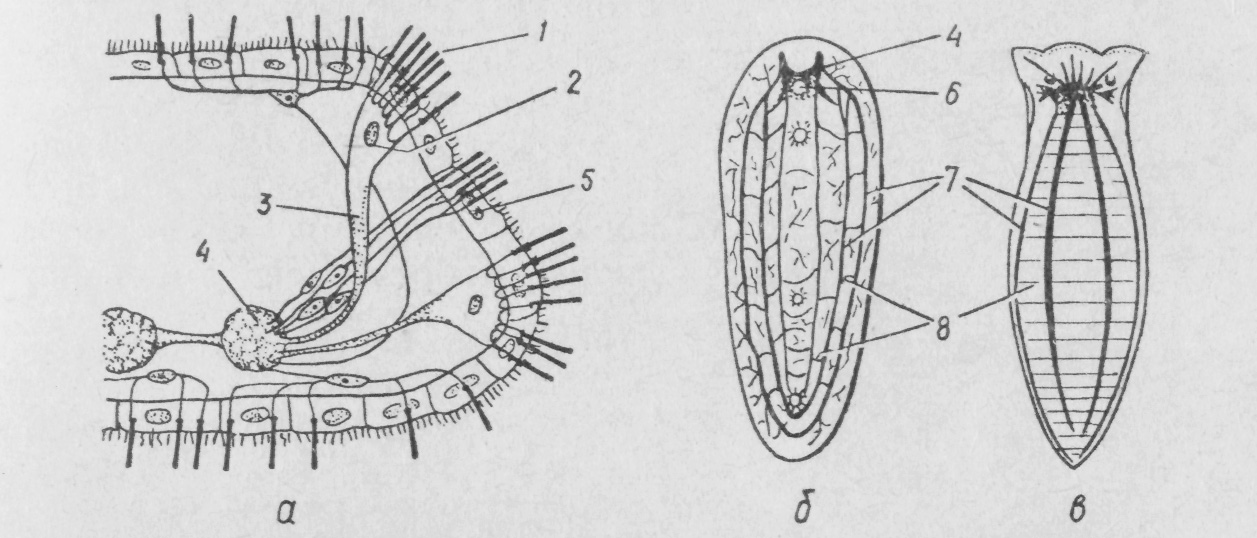
Подальша концентрація нервової системи призводить до того, що кількість поздовжніх і поперечних стовбурів зменшується до трьох-чотирьох пар. Лише в найпримітивніших представників ряду безкишкових і гнатостомулід нервова система розташована серед клітин епітелію, як і в кишковопорожнинних. У більшості видів вона занурюється під покриви, в товщі епітелію залишаються лише чутливі нервові клітини.
У деяких турбеляцій ендонний мозок не розвивається, замість нього утворюється так званий ортогонний мозок із
розширених передніх кінців стовбурів ортогону та потовщеної передньої кільцевої ко-місури.
Мозковий ганглій турбеля-рій стає інтегруючим центром нервової системи, який координує роботу всієї нервової
системи, обробляє інформацію від рецепторів і здійснює взаємодію між організмом і зовнішнім середовищем.
У турбелярій добре розвинені органи чуття. Механо- та хеморецептори представлені поодинокими або зібраними докупи сенсилами — чутливими клітинами, які мають довгі нерухомі війки або джгутики й відростки, що йдуть до мозкового ганглію. Хеморецептори концентруються в аурикулярних органах — ямках на передньому кінці тіла, вистелених миготливим епітелієм і сенсилами (див. рис. 131).
У багатьох турбелярій є зв'язаний із мозком орган рівноваги — статоцист, що має вигляд пухирця, всередині якого знаходиться одне або кілька вапнякових тілець — статолітів.
О
Рис.
133. Схема будови статевої системи
турбелярій (Tricladida):
/ —
яєчник; 2
—
жовтівники; 3 — яйцепровід; 4— сім'яники;
5
—
сім'япровід; 6
—
копулятивна сумка; 7 — копулятивний
орган; 8
— статева
клоака; 9
—
залозистий орган;
10
— статевий
отвір; // — видільна система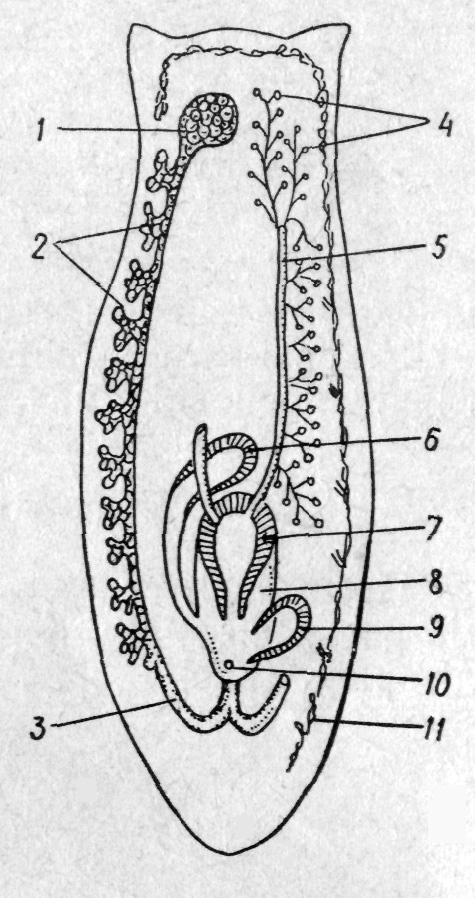 чі
турбелярій мають різну будову та
розташування. Як правило, в турбелярій
є пара очей, що містяться поблизу
мозкового ганглію. Однак у деяких великих
за розміром триклад і поліклад налічується
до кількох сотень очей, розташованих
по краях або на спинній стороні тіла.
Очі містяться в паренхімі, рідше — в
шкірному епітелії. Око найчастіше
складається з пігментованого бокала,
зверненого увігнутим боком до поверхні
тіла (рис. 132). У порожнину бокала входять
світлочутливі частини рецепторних
клітин, тіла яких лежать перед входом
до бокала, а їх протилежні
кінці утворюють довгі відростки, що
з'єднуються в пучок (зоровий нерв),
який іде до мозку. Отже, світлові промені
спочатку проходять через тіла рецепторних
клітин і лише потім потрапляють на їх
світлочутливі ділянки. Таке око
називається оберненим,
або інвертованим.
чі
турбелярій мають різну будову та
розташування. Як правило, в турбелярій
є пара очей, що містяться поблизу
мозкового ганглію. Однак у деяких великих
за розміром триклад і поліклад налічується
до кількох сотень очей, розташованих
по краях або на спинній стороні тіла.
Очі містяться в паренхімі, рідше — в
шкірному епітелії. Око найчастіше
складається з пігментованого бокала,
зверненого увігнутим боком до поверхні
тіла (рис. 132). У порожнину бокала входять
світлочутливі частини рецепторних
клітин, тіла яких лежать перед входом
до бокала, а їх протилежні
кінці утворюють довгі відростки, що
з'єднуються в пучок (зоровий нерв),
який іде до мозку. Отже, світлові промені
спочатку проходять через тіла рецепторних
клітин і лише потім потрапляють на їх
світлочутливі ділянки. Таке око
називається оберненим,
або інвертованим.
Г
Рис. 132. Схема
будови ока турбе-лярій:
/ —
світлочутлива частина рецепторних
клітин; 2
—
ядра клітин пігментного бокала; 3
—
пігментний бокал; 4
—
нервові відростки рецепторних клітин,
що утворюють зоровий нерв; 5 — ядра
рецепторних клітин
У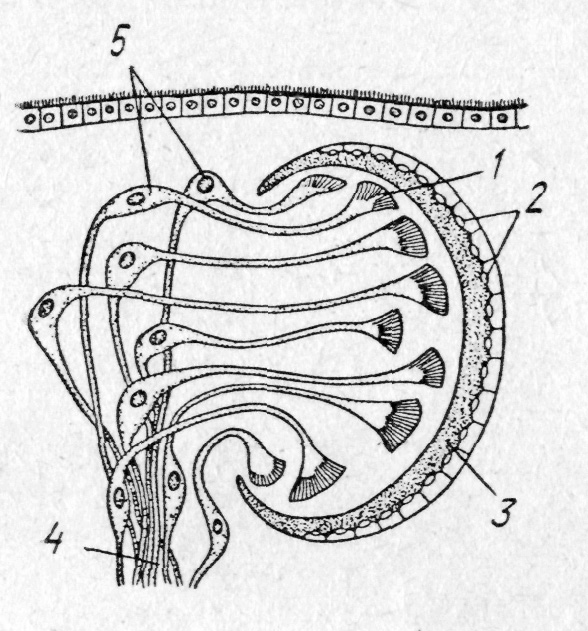 вищих турбелярій жіночі статеві залози
диференціюються на яєчники
та жовтівники,
що виробляють жовткові
клітини — видозмінені
ооцити, які втратили здатність до
запліднення й розвитку, а спеціалізуються
на утворенні жовтка. Вони входять до
складу складного яйця, й призначені для
живлення зародка,
що розвивається. Від яєчників відходять
яйцепроводи, в
які впадають протоки жовтівників.
Яйцепроводи зливаються позаду глотки
в одну трубку — піхву,
з нею часто зв'язана
копулятивна сумка, в
яку під час копуляції надходять
сперматозоїди (рис. 133).
вищих турбелярій жіночі статеві залози
диференціюються на яєчники
та жовтівники,
що виробляють жовткові
клітини — видозмінені
ооцити, які втратили здатність до
запліднення й розвитку, а спеціалізуються
на утворенні жовтка. Вони входять до
складу складного яйця, й призначені для
живлення зародка,
що розвивається. Від яєчників відходять
яйцепроводи, в
які впадають протоки жовтівників.
Яйцепроводи зливаються позаду глотки
в одну трубку — піхву,
з нею часто зв'язана
копулятивна сумка, в
яку під час копуляції надходять
сперматозоїди (рис. 133).
Ч
Рис. 134. Мюллерівська
личинка:
/—мозковий
ганглій; 2
—
мішкоподібний кишечник;
3
—рот; 4
— біляротові
лопаті; 5
— очі оловіча
статева система теж має складну будову.
Вона включає велику кількість сім'яників,
розсіяних у паренхімі,
від яких відходять сім'явивідні
канали, що з кожного
боку тіла впадають у один поздовжній
сім'япровід. Обидва
сім'япроводи ззаду зливаються, утворюючи
сім'я-випорскувальний
канал всередині
копулятивного органа,
який відкривається
в статеву клоаку.
оловіча
статева система теж має складну будову.
Вона включає велику кількість сім'яників,
розсіяних у паренхімі,
від яких відходять сім'явивідні
канали, що з кожного
боку тіла впадають у один поздовжній
сім'япровід. Обидва
сім'япроводи ззаду зливаються, утворюючи
сім'я-випорскувальний
канал всередині
копулятивного органа,
який відкривається
в статеву клоаку.
Запліднення турбелярій звичайно перехресне, для них характерний спіральний тип дробіння яйцеклітини.
Розвиток турбелярій прямий або з метаморфозом. У морських полікладід з яйця виходить так звана мюллерівська личинка. Вона має мішкоподібне тіло, на середині якого є вісім лопатей, облямованих війками; її кишечник не розгалужений (рис. 134). Личинка веде планктонний спосіб життя й лише після перетворення опускається на дно.
У переважної більшості війчастих червів спостерігається лише статеве розмноження. Разом із тим багато прісноводних трикладід, катенулід, макростомід розмножуються ще й вегетативно, шляхом поперечного поділу.
У класі турбелярій виділяють більше десяти рядів. Розглянемо основні з них.