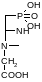Макроэргические молекулы
К макроэргическим относятся соединения, при гидролизе которых выделяется энергия не менее 7 ккал/моль. Это число — условная единица, означающая всего лишь некий уровень отсчета, согласно которому АТФ и несколько других соединений отличаются от остальных природных соединений. Соответственно название «макроэргический» употребляется для объединения соединений в группу веществ макроэргических и для указания их особой важности в переносе энергии в живой клетке.
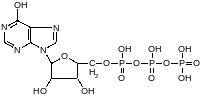
Нуклеозидтрифосфаты
Наиболее распространенными
высокоэнергетическими общими
промежуточными продуктами являются
нуклеозидтрифосфаты (НТФ), которые
могут передавать свою концевую
высокоэнергетическую фосфатную группу
любой из многочисленных органических
молекул - акцепторов (чаще всего энергия
образуется в виде АТФ). Особ енность
высокоэнергетических нуклеотидов
состоит в том, что они выступают в
качестве универсального источника
энергии для большого числа энергозависимых
реакций.
енность
высокоэнергетических нуклеотидов
состоит в том, что они выступают в
качестве универсального источника
энергии для большого числа энергозависимых
реакций.
Молекула АТФ состоит из аденилатной группы и трех остатков фосфорной группы. Значительная часть свободной энергии этой молекулы обусловлена взаимным электростатическим отталкиванием этих фосфатных остатков аналогично взаимному отталкиванию одноименно заряженных зарядов. Разрыв связей между остатками фосфорной кислоты сопровождается освобождением энергии. Соединением, играющим наиболее важную роль в клеточной энергетике, является АТФ, так как:
Химическая энергия запасается путем образования АТФ, сопряженного с катаболическими реакциями расщепления.
Затем химически энергия утилизируется путем расщепления АТФ, сопряженного с эндэргическими реакциями синтеза в ходе анаболизма и других процессов, требующих затрат энергии, например активного транспорта и сокращения мышц.
Гидролиз АТФ — термодинамическая движущая сила процессов, которые сами по себе являются термодинамически невыгодными.
АТФ и другие нуклеозидтрифосфаты ответственны за перенос энергии во многих сопряженных реакциях. АТФ — постоянный источник энергии для клетки. Он мобилен и может доставлять химическую энергию в любую часть клетки. Когда клетка нуждается в энергии, единственное, что потребуется для ее получения, — это гидролиз АТФ. АДФ может быть рефосфорилирован в АТФ в результате дыхательной активности или за счет другого высокоэнергетического соединения, например, креатинфосфата, присутствующего в мышечных клетках. Если весь АДФ мышечной ткани превращается в АТФ, то фосфат от АТФ переносится на креатин с образованием креатинфосфата. При этом вновь появляется некоторое количество АДФ, который может, присоединив фосфат, образовать АТФ. При понижении уровня АТФ происходит обратный процесс: фосфат переносится от креатинфосфата на АДФ, и запасы АТФ восстанавливаются.
Таким образом, АТФ играет важную метаболическую роль благодаря своему центральному положению в клеточной активности. Он действует как связующее клеточное звено между дыханием и процессами, требующими затраты энергии. При этом его высокоэнергетические фосфатные группы непрерывно отщепляются и замещаются новыми.
Аргининфосфат и креатинфосфат выполняют роль своеобразных аккумуляторов химической энергии, которые используются для быстрого фосфорилирования АТФ во время энергичного мышечного сокращения. Их называют фосфагенами.
Ацилфосфаты — макроэргические соединения с ангидридной связью, в которых карбонильный атом углерода ацильной группы особенно легко участвует в реакции с нуклеофилами. Значение G = -12,8 ккал/моль.
Например, глицеро-1,3-дифосфат + НОН глицеро-3-фосфат + Н3РО4.
Тиоэфиры играют очень важную роль в метаболизме в качестве метаболически активной формы ацильной группы. В природе основными тиолсодержащими соединениями являются: кофермент А, липоевая кислота, белки с -SH группой:
ацетил-КоА + НОН ацетат + HSKoA;
G = -7,37 ккал/моль.
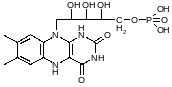
Восстановленные формы НАДН + Н+ и НАДФН + Н+, будучи синтезированными в клетке, затем вновь окисляются, при этом происходит перенос электронов на кислород. Этот способ используется в качестве основного способа, посредством которого клетка превращает химическую энергию поступивших извне питательных веществ в утилизируемую метаболическую энергию.
С тандартная
энергия гидролиза некоторых соединений
тандартная
энергия гидролиза некоторых соединений
G, ккал/моль
Нуклеозидтрифосфаты —7,3
1,3-Дифосфоглицерат —11,8
Фосфоенолпируват —14,8
Ацилтиоэфиры —7,37
Креатинфосфат —10,3
Восстановленные эквиваленты (при окислении):
НАДН + Н+; НАДФН + Н+ —52,6
Структура макроэргических соединений
НТФ, например ГТФ:
1,3-Дифосфоглицерат:
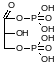
Фосфоенолпируват:
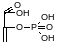
Ацилтиоэфиры:
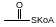
Восстановленные эквиваленты, например ФМН-Н2:
Креатинфосфат: