

5.5. Структура тРнк
Молекули тРНК являють собою полінуклеотидний ланцюг, що містить від 75 до 90 нуклеотидів. В організмі тРНК виконують дві основні функції: акцепторну й адапторну. Акцепторна функція полягає в тому, що аміноацил-тРНК-синтетаза приєднує амінокислоту до відповідної тРНК. Комплекс тРНК – амінокислота виступає в ролі адаптора (посередника) між мРНК і поліпептидним ланцюгом, що синтезується. тРНК є однією з найбільш вивчених макромолекул. Проведено аналіз первинної структури близько 200 молекул тРНК. Характерною її особливістю є наявність в її складі приблизно 10 % мінорних нуклеотидів. тРНК утворюють вторинну структуру, яка нагадує лист конюшини (рис. 5.13).
Кожна із чотирьох ділянок “листа конюшини” складається із чотирьох–семи комплементарних пар, які утворюють подвійні спіралі. Існують ділянки тРНК Д, Т, акцепторна й антикодонова. Д- і Т-ділянки названо так тому, що вони містять відповідно дигідроуридин і тимідин. Кількість нуклеотидів у ділянках і петлях загалом є сталою, за виключенням варіабельних петель, які в більшості тРНК містять чотири–п’ять нуклеотидів (клас І), але деякі – від 13 до 21 нуклеотиду (клас ІІ).
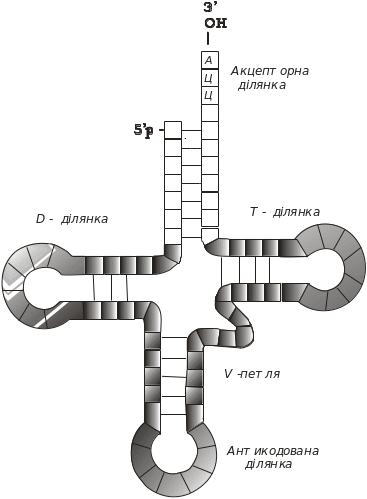
Рис. 5.13. Вторинна структура тРНК
Третинна структура тРНК. На початку 70-х рр. ХХ ст. у результаті рентгеноструктурних досліджень кристалів тРНК було встановлено, що вторинна структура листка конюшини тРНК згорнута у третинну структуру, яка нагадує літеру Г (рис. 5.14).
Рентгеноструктурні дослідження показали, що всі тРНК мають більш-менш однакову Г-подібну структуру. Два кінці літери Г, утворені акцепторною й антикодоновою ділянками, мають довжину приблизно по 7 нм і розміщені на відстані один від одного приблизно на 8 нм. Товщина цих ділянок становить близько 2 нм, тобто відповідає діаметру подвійної спіралі А-РНК. Молекула тРНК має псевдосиметрію другого порядку. Вторинна й третинна структури тРНК стабілізовані горизонтальними взаємодіями між комплементарними парами, а також вертикальними стекінг-взаємодіями. У тРНК виявлено особливий вид стекінгу, коли основи з однієї ділянки ланцюга інтеркалюють (вбудовуються) між двома основами іншої. При біосинтезі білка зв’язана з рибосомою аміноацил-тРНК має впізнати “свій” кодон на мРНК. Це забезпечується шляхом утворення подвійної мініспіралі між кодоновим й антикодоновим триплетами.
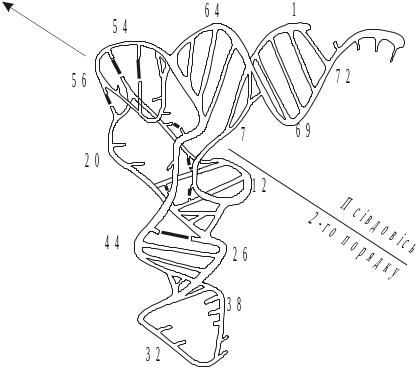
Рис. 5.14. Схематичне зобоаження просторової структури тРНКPhe із дріжджів
5.6. Рівні компактизації днк
Молекули ДНК в еукаріотів є значно більшими й місятться в кількох хромосомах, які мають складну організацію. У клітинах людини присутні 46 хромосом з ДНК, довжина яких коливається від 4 до 8 см (сумарна довжина хромосомних ДНК людини становить приблизно 2 м). Характерною особливістю еукаріотичної ДНК є високий рівень її компактизації. У клітинах еукаріотів хромосомна ДНК у комплексі з гістонами утворює хроматин (рис. 5.15). Спочатку вважалося, що ДНК у хроматині рівномірно вкрита шаром гістонів, але за допомогою електронної мікроскопії встановлено, що це не відповідає дійсності.
Гістони – це білки, які містять у значній кількості основні амінокислотні залишки. С-кінцеві ділянки гістонів Н2А, Н2В, Н3 та Н4 є гідрофобними, а N-кінцеві – гідрофільними (основними). Вважається, що гідрофобні ділянки гістонів мають виражену вторинну (спіральну) структуру. Гістон Н1 за своїми властивостями відрізняється від інших гістонів; він має іншу молекулярну масу й зв’язується безпосередньо з ДНК, тоді як інші чотири гістони утворюють октамер, який містить по дві молекули гістону кожного типу. Молекула ДНК обвивається навколо цього октамеру, формуючи 1 ¾ лівих надвитків. Молекулярні властивості гістонів наведено в табл. 5.3.
Таблиця 5.3. Молекулярна маса й вміст амінокислотних залишків у гістонах
|
|
Кількість амінокислотних залишків |
|||
Гістон |
Молекулярна маса |
Усього |
Основних |
Lys |
Arg |
Н1 |
22 000 |
220 |
65 |
62 |
3 |
Н2А |
14 000 |
129 |
30 |
14 |
12 |
Н2В |
13 800 |
125 |
30 |
20 |
8 |
Н3 |
15 300 |
135 |
33 |
13 |
18 |
Н4 |
11 300 |
102 |
26 |
11 |
14 |
Lys, Arg i His.
Гістоновий октамер за формою нагадує серце; центральну та нижню його частини утворюють тетрамер Н32, Н42, а верхні виступи – два димери (Н2А, Н2В). Октамер, можливо, стабілізується гідрофобними взаємодіями між С-кінцевими ділянками гістонів, а багаті на аргінінові й лізинові залишки N-кінцеві ділянки розміщуються на поверхні октамеру й зв’язуються з ДНК, яка обвиває гістоновий кор. Нуклеосома, що складається з гістонового октамеру й навитих на нього лівих (1,75) витків ДНК має симетрію другого порядку.
Якщо ДНК у нуклеосомі перебуває в В- формі (має діаметр 2 нм), то при діаметрі нуклеосомної кор-частинки 11 нм на один виток суперспіралі ДНК припадає 80 пар основ (рис. 5.15, 5.16).
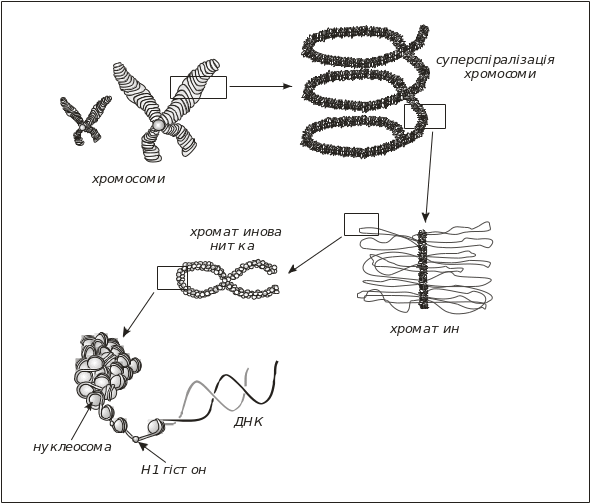
Рис. 5.15. Рівні компактизації ДНК
При закручуванні ДНК навколо гістонових октамерів відбувається локальна деформація подвійної спіралі. Вважається, що взаємне відштовхування між негативно зарядженими фосфатними групами сприяє тому, що подвійна спіраль ДНК існує у вигляді жорсткого стрижня. Але теоретичні розрахунки показують, що молекулу ДНК можна плавно обвити навколо гістонового кору. На користь цього свідчить також те, що основні амінокислоти в N-кінцевих ділянках гістонів частково нейтралізовуватимуть негативні заряди фосфатних груп ДНК. На рис. 5.16 наведено фрагмент ДНК, який утворює 1,75 витка навколо гістонового октамеру.
Усі гістони, за виключенням гістону Н1, зв’язуються з ДНК таким чином, що нейтралізують від’ємні заряди ДНК лише з одного боку подвійної спіралі. З протилежного боку відштовхування зарядів залишається попереднім, що сприяє вигину й обвиванню ДНК навколо гістонового кору.
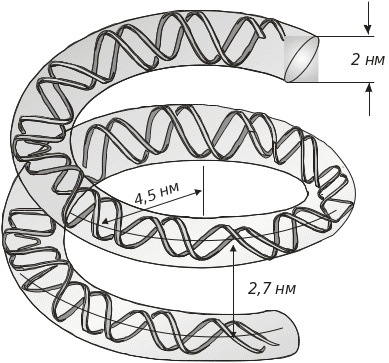
Рис. 5.16. Форма й розміри ДНК, яка обвиває гістоновий октамер. Крок суперспіралі 2,7 нм. На один супервиток припадає 80 пар основ ДНК